
Wie ist das Leben entstanden? Theorien und Experimente
Ein Beitrag von: Renate Ell - Stand: 02.11.2023
https://www.br.de/radio/bayern2/sendungen/iq-wissenschaft-und-forschung/wie-ist-das-leben-entstanden-theorien-und-experimente-100.html
https://www.preprints.org/manuscript/202201.0364/v2
https://www.wiko-greifswald.de/mediathek/beitrag/n/aus-dem-sternengas-zur-ersten-zelle-der-ursprung-des-lebens-in-der-kontinentalen-kruste-196975/
Zum Einstieg:
Aus dem Sternengas zur ersten Zelle – Der Ursprung des Lebens in der kontinentalen Kruste 28.05.2024
Öffentlicher Abendvortrag im Rahmen der Vortragsreihe „Ursprünge entdecken: Eine Reise von den Anfängen der Erde bis zur Entstehung der Menschheit“ von Professor Dr. Ulrich Schreiber (Duisburg-Essen)
https://www.wiko-greifswald.de/mediathek/beitrag/n/aus-dem-sternengas-zur-ersten-zelle-der-ursprung-des-lebens-in-der-kontinentalen-kruste-196975/
Um die Ursache von Ebbe und Flut zu erforschen hilft es nicht, in den Ozean zu schauen.
Die genauest mögliche Untersuchung einer Rakete oder eines Flugzeugs gibt keinen einzigen Hinweis auf die Entwicklung der ersten menschlichen Werkzeuge, wie Faustkeil oder Messer. Und doch bilden diese die Grundlage für die technische Revolution.

Wie und wo entstand das Leben?
Die kontinentale Erdkruste als Modellraum für die Bildung der ersten Zelle
Ulrich C. Schreiber; Bonn, Nov. 2022
Mai 2024
Update 16.12.2025 (überarbeitet)
Zusammenfassung
Die Frage nach der Entstehung des Lebens mit der Speicherung biochemischer Information beschreibt ein Problem, das trotz unterschiedlichster Ansätze in der naturwissenschaftlichen Forschung bislang ungelöst ist. Als Ursache kann gelten, dass das hierfür notwendige Environment nicht erkannt wurde. Aus geologischer Sicht bietet die geschützte Umgebung der kontinentalen Kruste ideale Bedingungen. Mit diesem Umfeld eröffnen sich völlig neue Möglichkeiten zur Identifizierung von Prozessen, die für die Entstehung der ersten Zelle entscheidend gewesen sein können. Vor diesem Hintergrund wurde die Theorie entwickelt, dass zelluläres Leben mit einem druckgesteuerten Zyklus im Grenzbereich überkritischer Gase begann. Die begleitende Speicherung von Peptidsequenzen in einer RNA erfolgte in einer Krustentiefe von etwa 400 bis 1000 Metern in unterschiedlich großen Kavitäten. Der Zyklus war in einem offenen System möglich, das an gasdurchlässige tektonische Bruchzonen mit einem hohen Anteil an CO₂ und N₂ aus dem Erdmantel gebunden war. Die Bildung von Peptiden und Vesikeln in überkritischem CO₂/N₂, sowie die chemische Evolution von Peptiden sind für die obere kontinentale Kruste bereits nachgewiesen. Weitergehende Überlegungen umfassen die Wechselwirkungen von Vesikeln mit katalytischen Peptiden und die Bildung einer Proto-tRNA. Hydrophobe Aminosäuren reichern sich in der Vesikelmembran an, wobei ihre Position in der Membran vom Grad ihrer Hydrophobizität abhängt. Mit Adenin an der Spitze des Akzeptorarms der Proto-tRNA kann die Membran durchdrungen werden, sodass eine Verknüpfung mit einer Aminosäure im Kontakt möglich wird. Die Eindringtiefe wird durch die Hydrophobizität des gegenüberliegenden Anticodons gesteuert. Durch Trennung der Proto-tRNAs in Einzelstränge werden Templates gebildet, die eine Verknüpfung von Aminosäuren zu kurzen Peptiden ermöglichen. Durch Zerfall der Vesikel bei jedem Zyklus werden die Reaktionsprodukte wieder in die Kavitäten abgegeben. Hier reichern sie sich an, wodurch mit jedem neuen Zyklus die Vesikel eine komplexere Mischung aufnehmen. Ab einem bestimmten Zeitpunkt war die Molekülzusammensetzung für die autonome Entwicklung der ersten Zelle erreicht.
Die alles umfassende Frage zur Entstehung des Lebens
Wie konnte aus anorganischen Stoffen, aus Gasen, Wasser und Gesteinen eine organische Chemie entstehen, aus der sich ein biologisches Informationssystem entwickelte? Die Speicherung von Information über die Baupläne der Zellen, mit all ihren biochemischen Instrumenten und abgestimmten Reaktionsabläufen stellt die absolute Grundlage des Lebens dar. Und wie schaffte es das Leben, sich gegen die allgegenwärtige Zunahme der Entropie zu behaupten und geordnete Strukturen aufzubauen? Diese Fragen gehören zu den größten der Wissenschaft, wenn man von denen der eigentlichen Entstehung des Weltalls absieht.
Das Wichtigste zuerst – Das Environment
In den vergangenen Jahren haben wir an der Universität Duisburg-Essen zusammen mit dem Physikochemiker Prof. Dr. Christian Mayer ein Modell entwickelt, das eine naturwissenschaftliche Erklärung für die ersten Schritte auf dem Weg zum Leben in der jungen kontinentalen Kruste bietet [1]. Das Modell wird gestützt durch Experimente mit einer Hochdruckzelle (Abb. 1), die realistische Verhältnisse, wie sie auch heute noch in der Kruste existieren, simuliert. Es konnte bereits ein weitergehendes Verständnis über die Molekülherkunft, ihre Aufkonzentrierung, die Fragen nach Energie und Entropie sowie die der Membranbildung erreicht werden [2].

Abb. 1 links: Hochdruckanlage, rechts: Ausschnitt mit isolierter Druckzelle (50 ml)
Worum geht es?
Die Verhältnisse der jungen Erde und die Bedingungen für die Entstehung des Lebens liegen aufgrund der großen zeitlichen Distanz zu heute weitgehend im Dunkeln. Fehlende Randbedingungen verhindern eine klare Eingrenzung der möglichen Prozesse, sodass in einer Vielzahl von Vorläufermodellen immer nur eng begrenzte Aussagen zu einzelnen Reaktionen getroffen werden konnten. Hierbei wurden als mögliche Environments alle Lokalitäten auf der Erdoberfläche, von der Tiefsee über vereiste Regionen bis zu flachen Tümpeln diskutiert. Aufgrund von Kritiken an den Modellen und mangels plausibler Alternativen gab es in der letzten Zeit Vorschläge, extraterrestrische Regionen wie den Mars oder Meteorite im Weltraum insgesamt mit einzubeziehen.

Abb. 2 Grundgebirgsaufschluss in Nord-Norwegen auf dem Weg nach Kirkenes, nahe der Grenze zu Finnland - Störungen und Risse im Gesteinsverband sind durch granitische Magmen (rosafarbene Gänge) oder hydrothermal gebildete Quarze (weiße Gänge) ausgefüllt.
Die kontinentale Kruste, die vor 4 Mia. Jahren immerhin schon auf 25% der heutigen Größe angewachsen war [3], wurde hingegen erst von der Essener Forschergruppe näher betrachtet [1]. Sie unterscheidet sich von der ozeanischen Kruste durch eine geringere Dichte und Temperatur, größere Mächtigkeit sowie durch eine vielfältigere mineralogische und chemische Zusammensetzung. Innerhalb der kontinentalen Kruste liegen bei genauem Hinsehen fast schon optimale Verhältnisse für die Startphase des Lebens vor. Durch die Analyse der dortigen Rahmenbedingungen konnte eine Vielzahl von neuen Ansätzen gefunden werden, die helfen, Probleme zu lösen, die in anderen Modellen als unlösbar gelten. Ausgangspunkt sind tiefreichende tektonische Störungszonen, die Kontakt zum Erdmantel haben. Sie sind vergleichbar mit der San-Andreas Störung oder dem Bayerischen Pfahl im Bayerischen Wald, der mehr als 200 km lang und als Störungszone seit mehr als 250 Millionen Jahre aktiv ist. Von der Tiefe steigen, wie zum Beispiel auch heute noch in der Eifel, Wasser, CO2 und andere Gase auf, die alle erforderlichen Stoffe für die Bildung organisch-chemischer Moleküle enthalten (Abb. 3).
Eines der kräftigsten Argumente diese Region hinsichtlich der Entstehung des Lebens genauer zu untersuchen ist das CO2, das in einer Tiefe von ca. 800 bis 1000 Metern einen Phasenwechsel von überkritischem Gas (ükCO2, wie es in der Tiefe vorliegt) zu gasförmigem CO2 (unterkritisch, gCO2, für den oberen Bereich) vollzieht (kritischer Punkt des reinen Gases bei 31 °C und 73,8 bar). Im überkritischen Zustand nimmt ükCO2 Eigenschaften sowohl von einer Flüssigkeit, in der Stoffe gelöst werden, als auch einer Gasphase an. Im ükCO2 können viele chemische Reaktionen ablaufen, die im Wasser nicht möglich sind. Es wirkt wie ein organisches Lösungsmittel und erweitert die Zahl der möglichen chemischen Bildungsreaktionen erheblich. Mit Wasser bildet es darüber hinaus Grenzflächen, an denen spezielle Reaktionen stattfinden können.
Durch Druckabfall geht in der Grenzregion von ca. 1000 m bis 800 m Tiefe das ükCO2 in gCO2 über (der Druck variiert hier durch die Anzahl der Gasblasen in der übestehenden Wassersäule). In dessen Folge enstehen Vesikel mit der Ausbildung einer Doppelschicht-Membran, die als Vorläufer der Zelle angesehen werden können (s.u.). Neben CO2 ist Stickstoff (N2) ein weiteres Gas, das in großer Menge aus dem Mantel in die Atmosphäre gefördert wurde und in den Aufstiegskanälen der Störungszonen in Mischungen mit CO2 oder auch in reiner Form aufgetreten sein muss. Die physikalischen Daten (kritischer Punkt des reinen N2 Gases bei −146.9 °C und 33,96 bar) zeigen, dass der überkritische Zustand bereits bei etwa 350 m Tiefe in einer offenen Wassersäule eintritt (wieder in Abhängigkeit von der Anzahl der Gasblasen im überstehenden Wasser). Das bringt für einige chemische Reaktionen Vorteile, da der pH-Wert höher liegt und eher den neutralen Bereich abbildet.
Molekül-Fossilien aus der Natur
Im Labor lassen sich bereits grundlegende Schritte auf dem Weg zu einer Zelle belegen [4]. Hierzu gehören die Bildung von Vesikeln, die als erste zellähnliche Strukturen gelten können, sowie die Verknüpfung von Aminosäuren zu längeren Ketten, die die Basis für die Entstehung komplexer Moleküle wie Proteine und Enzyme sind. Besonders attraktiv für das Modell ist die Tatsache, dass die Bedingungen für die Entstehung des Lebens in hydrothermalen Ganggesteinen aus der Frühzeit der Erde zum Teil dokumentiert wurden. In winzigen Flüssigkeitseinschlüssen, wie sie in archaischen Gangquarzen Australiens und aus entsprechenden Geröllen in Konglomeraten (Abb. 4) von uns gefunden wurden (Quarze kristallisieren unter hydrothermalen Bedingungen in Störungszonen), verbirgt sich eine Vielzahl organischer Stoffe aus dieser Zeit [5]. Sie wurden vor Milliarden Jahren während der Kristallbildung eingeschlossen und so konserviert. Sie helfen uns, die Bedingungen für die Laborversuche der Wirklichkeit anzunähern. Die aufwendige Analytik gelang durch Zusammenarbeit der Labore aus Heidelberg (AGs Heinz F. Schöler und Frank Keppler mit Ines Mulder, Tobias Sattler und Markus Greule) und Essen (Christian Mayer mit Maria Davila Garvin und AG Oliver J. Schmitz mit Pia Rosendahl, Amela Bronja und Yildiz Grossmann).

Abb. 3 Schematisches Blockprofil der frühen kontinentalen Kruste. Die durchschnittliche Mächtigkeit beträgt heute 30 km. Sie ist hier geringer angenommen. Gleichzeitig muss die Temperatur höher gelegen haben, konnte aber durch artesische Wässer im oberen Bereich gekühlt sein.


Abb. 4 Konglomerat in den Jack Hills, Westaustralien, von dem Gerölle hydrothermaler Quarze analysiert wurden. In dem Gestein sind die bisher ältesten Zirkone gefunden worden (> 4 Mia. J.).
Selbst in fast rezenten Mineraleinschlüssen finden sich in entsprechendem Environment gleiche Molekülreihen (Aldehyde), die anorganisch aus den Gasbestandteilen der Kruste gebildet wurden. So konnten aus den CO2-führenden Spalten des jungen Wehr-Vulkans (westlich vom Laacher- See, Eifel) aus fast 1000 m Tiefe Kalzite aus einem Bohrkern gewonnen werden (Abb. 5), die eine vergleichbare Chemie der archaischen Quarze aus Australien zeigt. Die Analysen wurden maßgeblich in Zusammenarbeit mit
Prof. Oliver J. Schmitz in der Fakultät für Chemie der Universität durchgeführt [6].

Abb. 5 Mit Kalziten mineralisierte Kluft im Bohrkern aus dem Wehrer Kessel (ca. 960 m Tiefe), letzte Eruption vor ca. 150.000 Jahren, Rissbildung im Gestein durch Vulkanotektonik, nachträglich mineralisiert mit Calzitmineralen, in denen es Flüssigkeitseinschlüsse mit komplexer organischer Chemie gibt. Foto: Prof. Dr. O.J. Schmitz
Eine besondere Herausforderung stellt aber die Erforschung der Entwicklung des biologischen Informationssystems in der DNA bzw. in dem vermuteten Vorläufer der RNA dar. Die physikochemischen Voraussetzungen in der kontinentalen Kruste scheinen auch für diesen Prozess günstig gewesen zu sein. Erste Ansätze für eine Lösung zeichnen sich bereits ab.
Die wichtigsten Grundlagen zur Entstehung des Lebens
Da es zu Beginn noch keine molekularen Werkzeuge wie Enzyme gab, müssen die ersten Schritte zum Leben auf rein physikalische bzw. physikochemische Weise abgelaufen sein. So lässt sich die Bildung einiger der Komponenten, die für die Zelle benötigt werden, durch Prozesse erklären, die heute in der technischen Chemie täglich eingesetzt werden. Hierzu gehören Verfahren der Fischer/Tropsch-Synthese, durch die langkettige Kohlenwasserstoffe gewonnen werden oder auch die Haber/Bosch-Synthese, durch die Ammoniak (NH3) aus N2 und H2 produziert wird. Die Druck/Temperatur-Bedingungen, die hierfür notwendig sind, liegen in der Kruste genau so vor, wie die erforderlichen Rohstoffe. Weiterhin ist zu überlegen, welche Bedingungen und Prozesse insgesamt vorhanden waren bzw. abgelaufen sind, die in einem Entstehungsumfeld gestaltend auf die Entwicklung des Lebens eingewirkt haben.
Die Entropiezunahme als Voraussetzung für die Entstehung des Lebens
- Durch die Verknüpfung von Molekülen, z.B. von zwei Aminosäuren, entsteht mehr Ordnung, als vorher in dem betrachteten System vorhanden war. Die chemische Reaktion (z.B. eine Kondensationsreaktion) erfolgt nur, wenn gleichzeitig ein höheres Maß an Unordnung gebildet wird, also die Entropie insgesamt zunimmt. Es könnte z.B. im Reagenzglas durch eine Reaktion Wärme entstehen, die nach außen abgegeben wird, wodurch die Entropie erhöht wird. Die Entropiezunahme (häufig als Zunahme der Unordnung beschrieben) muss folglich im gesamten Verlauf der Lebensentwicklung während der Reaktionen gewährleistet gewesen sein.
Der Übergang von überkritischem CO2 (ükCO2) zu Gas (gCO2) in ca. 800 - 1000 m Tiefe ist mit einer starken Zunahme der Entropie verbunden.
- Für den Ablauf chemischer Reaktionen ist im Normalfall Energie notwendig, die in erforderlichem Maße zur Verfügung stehen musste.
Zur Verfügung standen in der Kruste Wärmeenergie, potentielle Energie (aus dem Aufstieg von Gasen) und chemische Energie (zum Teil auch elektrische Energie durch piezoelektrischen Spannungsaufbau infolge seismischer Aktivität). Zusätzlich gab es einen nicht zu vernachlässigenden Strahlungsanteil infolge Zerfall radioaktiver Elemente in den Mineralen.
- Das Umgebungsmilieu (Environment) muss über sehr lange Zeiträume stabil und vor UV-Strahlung, Sonnenwind, Verwitterung, Meteoriteneinschlägen oder auch vor Erosion geschützt gewesen sein. Notwendig waren Hunderte Millionen Jahre.
Störungszonen wie z.B. die San-Andreas Störung oder der Bayerische Pfahl erreichen diese Lebensdauer.
- Das System musste offen im Ungleichgewicht sein mit der Möglichkeit der Zu- und Abfuhr von Stoffen. Einerseits wurden Moleküle verbraucht, andererseits durften „unbrauchbare“ Moleküle oder Reaktionsprodukte das System nicht verstopfen (Problem der Teerbildung) [7].
Durch die aufsteigenden Gase in den Störungen wurden Wasser und Moleküle in Richtung Oberfläche transportiert.
- Es müssen zyklische Wechsel der physikalischen Bedingungen geherrscht haben. Zyklische Bedingungen führen eher zu einer chemischen Evolution als sporadische Ereignisse.
Durch Ausbrüche von Kaltwasser-Geysiren oder durch die damals noch wesentlich stärkeren Erdgezeiten (der Mond war deutlich näher an der Erde) gab es zyklische Druckschwankungen, die im Übergangsbereich von ükCO2 zu gCO2 zu einem ständigen Wechsel der Bedingungen geführt haben.
- Die Rohstoffe zur Bildung von organischen Molekülen sowie notwendige Elemente (Metalle, Halbmetalle) mussten in großer Menge vorgelegen haben.
Durch die Zufuhr von Gasen (CO2, N2, H2, H2S etc.) aus dem Mantel und der Alteration der Gesteine mit Freisetzung von Phosphat und Metallen/Halbmetallen gab es unbegrenzte Ressourcen.
- Die Bildung der verschiedenen organischen Moleküle erforderte unterschiedliche Druck- und Temperatur- (pT-) Bedingungen. Ein Transportsystem zum Zusammenführen der unterschiedlichen Molekülbausteine, die an verschieden Orten gebildet wurden, war eine der wichtigsten Voraussetzungen.
Durch die aufsteigenden Gase in den Störungen wurden Wasser und in unterschiedlichen Tiefen gebildete Moleküle in Richtung Oberfläche transportiert.
- Kondensationsreaktionen von z.B. Aminosäuren zu einem Peptid können nur stattfinden, wenn das bei der Reaktion entstehende Wassermolekül entfernt wird. Das ist im Wasser (z. B. der Zelle) nur mit Enzymen möglich, sodass in der Frühphase ein unpolares Lösungsmittel am besten hierfür geeignet war. Allerdings gab es noch keine Erdölchemie, die diese bereitstellen konnte.
In gasreichen Störungszonen liegt z.B. überkritisches CO2 oder überkritisches N2 als unpolares Lösungsmittel vor.
- Für die Bildung einer RNA (tRNA) als erste Informationsträger waren vermutlich anorganische Katalysatoren erforderlich. Tonminerale sind nach Laboruntersuchungen bereits als mögliche Katalysatoren diskutiert worden [8].
Tonminerale bilden sich nicht nur durch Verwitterung der Gesteine an der Erdoberfläche, sondern auch durch Alteration der gesteinsbildenden Minerale in der Erdkruste durch saure Wässer in den Störungen.
- Ein grundsätzliches Problem auf der jungen Erde war die Bildung einer großen Anzahl verschiedener organischer Moleküle. So auch in den Störungzonen der kontinentalen Erdkruste. Es herrschte quasi ein Molekül-Chaos, aus dem spezielle Moleküle noch nicht von „biochemischen Werkzeugen“ herausgefiltert werden konnten. Für eine Vorsortierung standen nur physikalische bzw. physikochemische Prozesse zur Verfügung:
>Strömungsprozesse in den wassergefüllten Störungen der kontinentalen Kruste durch aufsteigende ükGase (bzw. Gase in den obersten 800 Metern).<
>Oberflächeneffekte, wodurch es während des Aufstiegs zur Anlagerung spezieller Moleküle an wechselnden Mineraloberflächen (gesteinsbildende Minerale, Gangerze, wie z.B. Blei-Zink- oder Eisenschwefel-Erze, Quarz, Tonminerale) kommen konnte.<
>Flotation durch ükCO2/ükN2-Bläschen, die einerseits unpolare Moleküle direkt aufnahmen oder an der Bläschen-Oberfläche bestimmte Amphiphile anlagerten, die als Sammlermoleküle für z.B. bestimmte RNA-Moleküle fungieren konnten.<
Beispiel: Molekültrennung wie in technischen Verfahren der Chromatopraphie
In den Spalten der Störungszonen sind durch die Fluidströmungen alle Voraussetzungen für Trennvorgänge aus Gemischen organischer Moleküle gegeben. Die Verhältnisse gleichen chromatographischen Analyse- und Trennungsverfahren, bei denen es durch ein dynamisches Wechselspiel zwischen Lösemittel und fester Phase zu Trennungen unterschiedlichster Moleküle kommt. Die Risse und Bruchflächen in der kontinentalen Kruste haben Querschnitte von Kapillargröße bis in den zehner Zentimeter Bereich hinein. Hiermit sind sie vergleichbar mit Weiten, die in technischen Trennungsverfahren Verwendung finden. Besonders interessant ist der Trennungsprozess, wenn der Aggregatzustand der mobilen Phase überkritisch ist. Entsprechende Effekte in der kontinentalen Kruste sind direkt mit der überkritischen Flüssigkeitschromatographie (SFC, supercritical fluid chromatography) vergleichbar, die überwiegend mit überkritischem CO2 durchgeführt wird. Bei diesem Verfahren werden Kieselgele als feste Phase verwendet. Kieselgele sind typisch für tieferreichende Störungen. Aus ihnen wird das Gangmineral Quarz gebildet, das häufig die Bruchflächen auskleidet und die Grundlage der Quarzgänge bildet. Es gibt auch Verfahren wie den simulated moving bed (SMB) process, bei dem die feste Phase den zu trennenden Substanzen entgegenläuft. Dies wäre der Fall, wenn Kieselgele im Fluidstrom der Störungen mit nach oben transportiert werden. SMB Verfahren werden zum Beispiel in der pharmazeutischen Industrie genutzt, wenn Moleküle mit unterschiedlicher Chiralität in großen Mengen getrennt werden müssen. Auch dient das Verfahren zur Reinigung von Fructose im industriellen Maßstab. Hierin könnte die Ursache für die Verwendung von Ribose bei der Bildung der RNA in den hydrothermalen Spaltensystemen der Kruste begründet sein. Durch chromatographische Trennung von Gemischen unterschiedlichster Zucker wäre in diesem Fall die Ribose am stärksten angereichert worden, sodass sie am leichtesten verfügbar war.
Zone der Molekülanreicherung
In der Grenzzone zwischen 1000 m bis ca. 800 m findet ein Phasenwechsel der Gase vom überkritischen zum unterkritischen (gasförmigen) Zustand statt (bei reinen N2-Systemen entsprechend in geringerer Tiefe). Durch Druckschwankungen z.B. bei Kaltwassergeysirausbrüchen (CO2/N2 gesteuerte Ausbrüche) variiert die Tiefe des Übergangs zur unterkritischen Gasphase über mehrere 10er Meter. Die Folge ist ein Ausfallen der organischen Moleküle, weil das sich bildende unterkritische Gas keine Moleküle mehr gelöst halten kann. Hierdurch entwickelt sich ein Anreicherungshorizont, in dem die Konzentrationen der Stoffe so hoch werden, dass chemische Reaktionen begünstigt sind.

Abb. 6 Beispiel für die Verhältnisse in einer Störung. Die Pfeile zeigen auf Fallen für aufsteigende Gase, die dann gefüllt werden, wenn die Spalte in der Kruste mit Wasser gefüllt ist (links, Foto: Dr. Frederik Kirst). In größeren Tiefen unterhalb von ca. 800 m sammelt sich ükCO2 unter den Vorsprüngen. Hier können im überkritischen Gas unpolare Moleküle angereichert werden und miteinander reagieren (Skizze rechts).
RNA
Die Bildung einer RNA (als Vorläufer der DNA) gehört zu den wichtigsten Voraussetzungen für die Entwicklung eines organisch-chemischen Informationssystems. Die Bedingungen in den Störungszonen mit der Bereitstellung von organischen Basen, dem Zucker Ribose und Phosphat aus der Auflösung der Apatite waren hierfür günstig [9]. Allerdings wurden in diesem Umfeld nicht nur die Moleküle gebildet, die in der heutigen RNA vorliegen. Es gab sicherlich eine Vielzahl von verschiedenen Zuckern, die auch jeweils noch in den chiralen D- und L-Versionen auftraten. Die Alteration der Krustengesteine führte an verschiedenen Abschnitten der Störungsoberflächen zur Auskleidung mit Tonmineralen. Sie fungierten vermutlich als Katalysatoren, die die Verknüpfung von Nukleotiden zu längeren RNAs steuerten [8]. Diese RNAs konnten aufgrund unterschiedlicher Zucker oder auch Basen sehr verschieden sein. Es müssen daher Prozesse identifiziert werden, die ohne Hilfe heutiger biochemischer Katalysatoren zu dem RNA-Typ geführt haben, der das weitere Geschehen auf dem Weg zur ersten Zelle bestimmt hat. Eine Grundlage für die Ausrichtung in die heute vorliegende Version könnten die oben beschriebenen SFB- und SMC-Prozesse gewesen sein, die durch chromatographische Trennungsschritte das Angebot in Hinblick auf Qualität und Quantität der betreffenden Moleküle steuerten.
Bei der Bildung von längeren RNA-Strängen kommt es schnell zur Verknäuelung, da sich komplementäre Abschnitte zusammenlagern und teilweise Doppelstrangabschnitte bilden [10]. Eine Kopie, die durch Anlagerung freier Nukleotide erfolgen könnte, wird hierdurch unterbunden, ein Kernproblem der RNA-Welt. Durch Erhöhung der Temperatur werden Doppelstränge aber geschmolzen und stehen anschließend bei Abkühlung für eine Dopplung zur Verfügung. Die Verhältnisse in den Störungen bieten für den Vorgang des RNA-Kopierens optimale Bedingungen. Die Temperatur in 1000 m Tiefe könnte durch Zutritt von Oberflächenwasser bei etwas über 50 °C gelegen haben. Durch Übergang des ükCO2 zur Gasphase in Folge eines Geysirausbruchs bringt eine Abkühlung durch die Ausdehnung des Gases mit sich (Joule-Thompson Effekt).
Der umgekehrte Vorgang komprimiert das Gas, was mit einer Temperaturerhöhung verbunden ist. Hierbei können Werte deutlich über 60 °C erreicht werden, die notwendig sind, um RNA-Doppelstränge zu trennen. Zusätzliche Temperaturveränderungen ergeben sich durch aus der Tiefe nachströmendes heißeres Wasser und anschließend rücklaufendes kühleres von weiter oben nach Ende des Ausbruchs. Mit der Abkühlung beginnt die Phase der Anlagerung komplementärer Nukleotide. Sind genügend Bausteine verfügbar, können bei jedem Ausbruchszyklus RNA-Stränge kopiert werden. Gleiche Prozesse können mit überkritischem Stickstoff (ükN2) in geringeren Tiefen und entsprechend kühleren Hintergrundtemperaturen ablaufen.
Wenn die Verhältnisse in einem Environment für eine RNA-Bildung geeignet sind, ist nicht automatisch die Grundlage für die Entwicklung eines Informationssystems gegeben. RNA-Stränge, die sich bilden, besitzen unterschiedliche Längen und werden schnell durch Hydrolyse wieder in kurze Abschnitte aufgeteilt. Die Hydrolyse eines RNA-Strangs ist im sauren Milieu bei pH-Werten zwischen 3 und 4, wie sie auch in den Fluiden der Störungen in der Tiefe vorliegen, am geringsten [11], das heißt, hier liegt der größte Stabilitätsbereich vor. Es bleiben auch nach einer groben Vorsortierung immer noch sehr viele unterschiedliche Moleküle übrig, von denen nur bestimmte für die weiteren Schritte erforderlich sind. Das bedeutet, dass für jede Entstehung komplexerer Bausteine aus einfachen Ausgangsmolekülen, die sich im weiteren Verlauf im System behaupten, ein Vorteil für den Erhalt vorgelegen haben muss. Dahinter steht der Begriff der chemischen Evolution. Und die Ursachen sind immer physikochemische Gesetzmäßigkeiten, auf denen die Stabilität der Moleküle beruht.
Ab einer bestimmten Länge ist die Wahrscheinlichkeit hoch, dass in dem ersten und letzten Drittel des RNA-Strangs Abschnitte mit jeweils komplementären Basen vorliegen. Das bedeutet, dass bei einer Krümmung die Basen beider Drittel gegenüberliegen und wie bei einem DNA-Doppelstrang über eine Basenpaarung eine Verbindung eingehen (Adenin A verbindet sich mit Uracil U, Cytosin C mit Guanin G). Das mittlere Drittel bildet hierbei eine Schleife, die je nach Länge unterschiedliche Größen einnimmt. Bei ausreichendem Umfang sind die Nukleobasen alle nach innen gerichtet. Besitzt die Länge für eine Schleifenbildung eine kritische Größe, sodass nur eine Basenpaarung für die Fixierung einer Schleife (Loop) zur Verfügung steht, kann sich der Loop nur bilden, wenn drei der Basen nach außen zeigen (durch Drehung innerhalb des Strangs). Es entsteht so ein Anticodon, das mit drei komplementären Basen eine Verbindung eingehen kann. Diese Anticodon befindet sich heute an dem einen Ende der tRNA (Transfer-RNA), die in den Zellen, allerdings wesentlich komplexer, den Mittler zwischen einer spezifischen Aminosäure und eines Informationscodes bildet.
Wenn sich diese frühe RNA mit einem Anticodon als Proto-tRNA im weiteren Verlauf durchgesetzt hat, stellt sich die Frage, wie groß bzw. wie einfach sie insgesamt gebaut gewesen war. Die Komplexität von heute hatte sie auf keinen Fall. Sie muss eine minimale Stranglänge gehabt haben, mit einer gleichzeitig besonderen Ausstattung. Dahinter steckt die Überlegung, dass kurze RNA-Stränge häufiger gebildet werden können und stabiler sind, als lange. Hinzu kommt, dass ein einzelnes Basenpaar leichter für eine zum weiteren Fortbestand notwendige Kopie des Stranges getrennt werden kann, als eine größere Anzahl.
Neben dem Loop mit Anticodon und dem einzelnen Basenpaar fehlt noch ein wesentliches Teil, das ich oben mit `besonderer Ausstattung´ bezeichnet habe. Es ist ein einsträngiger Abschnitt am anderen Ende, an den eine Aminosäure angebunden werden kann (s.u.). Damit liegt ein Prototyp einer einfachen Proto-tRNA vor, der auf eine minimale Länge von 12 (oder alternativ 13) Nukleotiden kommt. Er ist in Abb. 7 dargestellt, mit lediglich einem Basenpaar.

Abb. 7 Mögliche Proto-tRNA: Kürzest möglicher RNA Strang mit Bildung lediglich eines Basenpaares und einem verlängerten Einzelstrang, dessen drei Basen immer C, C, A (Cytosin, Cytosin, Adenin) sind. In der Schleife sind drei Basen nach außen positioniert, die das Anticodon bilden (hier AAA; nach außen gerichtete Position aufgrund von Platzproblemen bei Verwendung des Zuckers Ribose). Eine Alternative mit 13 Nukleobasen wird weiter unten diskutiert.
Hierbei taucht sofort die Frage auf, warum die Ribose als Zucker mit 5 Kohlenstoffatomen (Pentose) den RNA-Strang aufbaut und nicht eine der anderen Pentosen oder gar der Hexosen, die Moleküle mit 6 C-Atomen bilden. Im Labor lassen sich derartige alternativen Nukleotidketten herstellen. Die verschiedenen Zucker müssen letztlich in den betrachteten Environments in einer Vielzahl vorgelegen haben, da ihre Bildung nicht selektiv auf nur einen Typ beschränkt ist. Und davon ausgehend gab es vermutlich eine Reihe anderer RNA-Strang-Typen, die andere Zucker eingebaut hatten, die ebenfalls chiral waren und somit zusätzlich zwei unterschiedliche Händigkeiten besaßen.
Welche Selektionsprozesse können jetzt zur Wahl der Ribose geführt haben? Sicherlich sind wieder Stabilitätskriterien oder allein das Mengenangebot in Betracht zu ziehen, das sich durch die oben dargestellten Trennverfahren ergeben haben kann. Aber eine andere Ursache könnte die entscheidende sein, die aus dem engen Loop des beschriebenen Proto-tRNA-Strangs resultiert. Die aus Platzgründen nach außen gerichteten drei Basen müssen in Kombination mit den Zuckermolekülen so angeordnet worden sein, dass sie als eine Einheit, als Dreierblock ablesbar waren. Jede andere Konstellation, die dies nicht gewährleistete, führte in der Entwicklung nicht weiter.
Jetzt muss verglichen werden, ob die alternativen Zucker, sowohl die anderen Pentosen als auch Hexosen und andere, eine Konfiguration der RNA-Schleife ergeben können, die einen nach außen gerichteten „ablesbaren“ Dreierblock beinhaltet. Eine Anordnung der Basen, die eine Ablesbarkeit beeinträchtigt oder unmöglich macht, setzt sich nicht durch (weil in der weiteren Entwicklung keine Information über enzymatisch unterstützende Katalysatoren erhalten bleibt). Wenn sich diese Überlegung bestätigt, wäre die Ribose aus rein sterischen Gründen (strukturell am besten geeignet, um ein Anti-Codon zu bilden) aus der Vielzahl der Zucker in Kombination mit den Basen und dem Phosphat selektiert worden.
Auffällig ist, dass alle tRNAs, die heute spezifisch mit einer Aminosäure beladen werden, immer den gleichen Kopplungsstrang der Basenreihung CCA besitzen (Abb. 7). Dieser Bindungsstrang muss sich in der frühesten Phase als so erfolgreich herausgestellt haben, dass er bis heute ausschließlich genutzt wird. Die Synthetasen, die heutigen komplexen Enzyme, die die spezifische Beladung auf den spezifisch zugehörigen tRNAs vornehmen, erkennen die inzwischen komplexer gebauten tRNAs anhand ihrer zusätzlicher Schleifen und den darin enthaltenen speziellen Basen, sodass die sehr wichtige genaue Zuordnung der zu verknüpfenden Aminosäure gewährleistet ist. Es bleibt die Frage, warum die Version mit dem Akzeptorarm CCA als vermutlich alleiniger Vertreter aus der Gruppe der RNAs mit mittlerer Länge aus dem Selektionsprozess hervorging.
Die Ursache kann in einem Vorteil gelegen haben, der durch eine spezielle Eigenschaft der Proto-tRNA bedingt war. Prinzipiell hätte jede Aminosäure an jeder Proto-tRNA anbinden können. Wo lag die Möglichkeit einer spezifischen Zuordnung, die für den Aufbau eines Informationsspeichers notwendig war? Die heutigen Verhältnisse lassen zumindest eine grobe Unterscheidung treffen. Es gibt zwei Klassen von Synthetasen, die die spezifische Beladung der tRNAs vornehmen. Die Klasse-I-Synthetase katalysiert hydrophobe Aminosäuren am 2´- OH Ende der endständigen Ribose (Abb. 8) und die Klasse-II-Synthetase katalysiert die hydrophilen Aminosäuren am 3´- OH Ende [13].
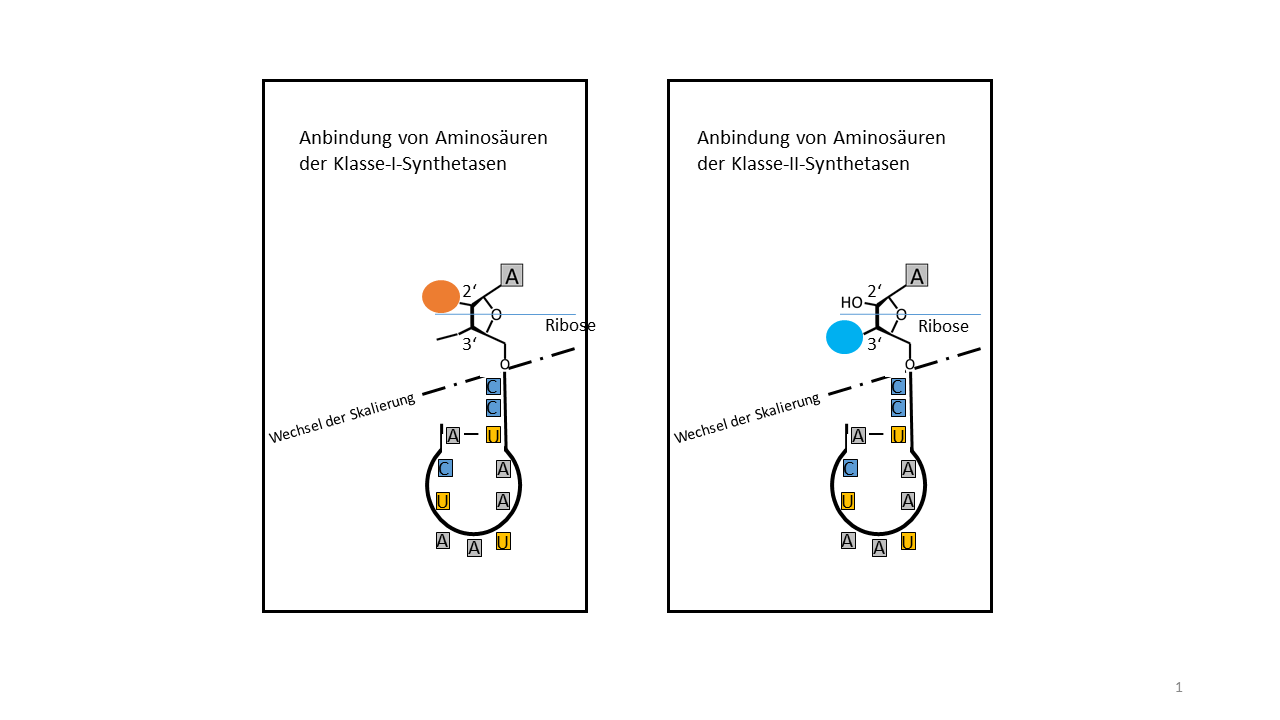
Abb. 8 Hydrophobe Aminosäuren werden mit der Klasse I tRNA Synthetasen an 2'-OH der Ribose und hydrophile Aminosäuren mit der Klasse II tRNA Synthetasen an 3‘-OH verknüpft. Die blaue Linie als Trennfläche zwischen einer hydrophoben (oben) und einer hydrophilen Umgebung (Bereich im Maßstab verzerrt) wäre eine mögliche Trennlinie für die beiden Verknüpfungsstellen in der Frühphase der Entwicklung.
Für die Anreicherung von bestimmten RNA-Molekülen können weiterhin Flotationsprozesse mit Hilfe von
Sammlermolekülen in Betracht gezogen werden. So können aufsteigende überkritische Gasblasen RNA-Moleküle in die Fällungszone transportiert haben. Hierzu sind Lipide (Amphiphile) in der Lage, die aus einem
hydrophilen Kopf und einem
hydrophoben Schwanzende bestehen (Abb. 9). Der Kopf ist in der Lage, sich mit einem RNA-Molekül (im Fall einer Proto-tRNA mit dem CCA-Ende ober weiteren Anknüpfungsstellen) zu verbinden (s. auch [14]). Während der hydrophobe Teil in eine überkritische Gasblase eingeschlossen werden kann, verbleibt der hydrophile Rest mit der RNA im Wasser. Die Gasblasen steigen auf und transportieren die Moleküle bis zu Gastaschen, in denen sie sich sammeln. Von dort oder auf direktem Weg gelangen sie bis zur Grenzzone, in der der überkritische Zustand in den unterkritischen wechselt. Während weitere RNA-Varianten ähnlich transportiert werden können, gehen hydrophobe organische Moleküle direkt in die Blasen über und steigen mit auf. In ca. 1000 Meter Tiefe werden mit dem Phasenwechsel durch Druckabnahme während eines Geysirausbruchs alle transportierten Moleküle im Wasser und an der Grenzfläche zum Gas angereichert.
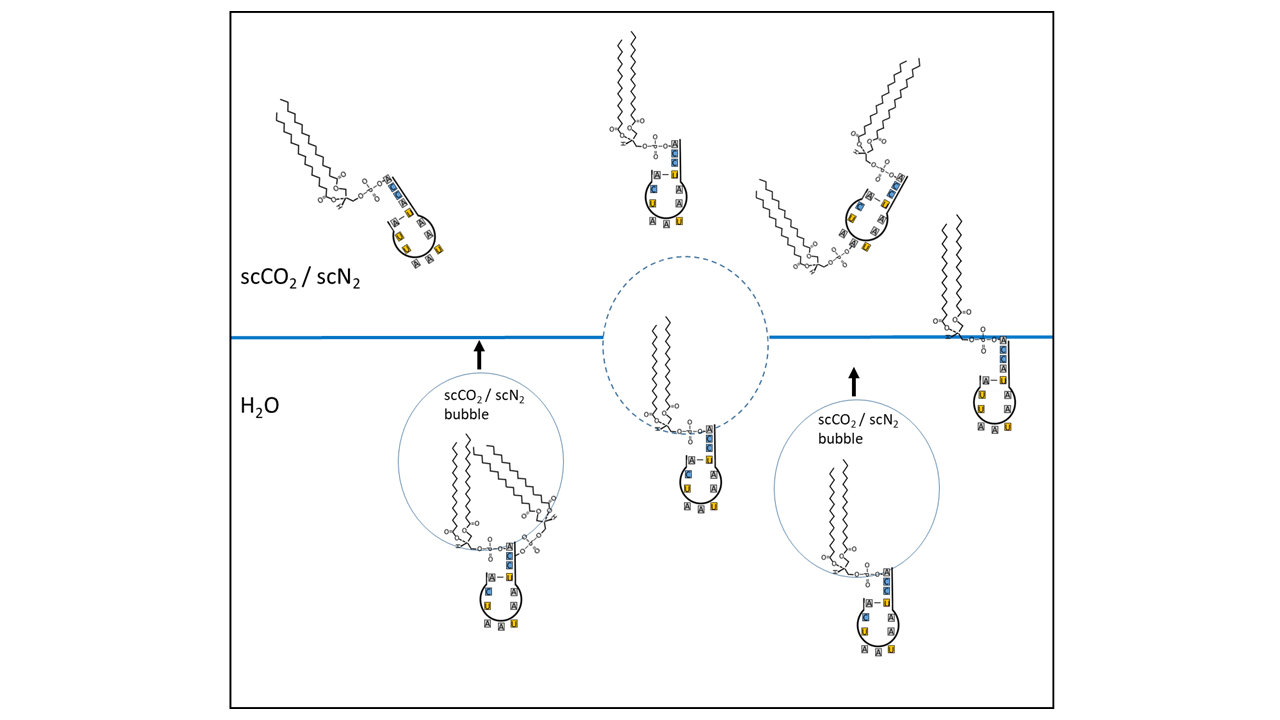
Abb. 9 Prinzipskizze eines Flotationsprozesses: Amphiphile Lipide sind am Akzeptorarm einer Proto-tRNA verknüpft. Der hydrophobe Teil ist in Blasen aus überkritischem CO2/N2 integriert, während der hydrophile Kopf mit der RNA im Wasser verbleibt. Die Blasen steigen auf und sammeln sich im Dachbereich einer Kavität. Ein Teil der Moleküle gelangt komplett in das ükCO2/N2 (sc = super critical, überkritisch) Reservoir, andere positionieren sich genau an der Wasseroberfläche. Dargestellt ist nur die postulierte Proto-tRNA als Schlüsselmolekül. Andere RNA-Varianten werden auf ähnliche Weise verknüpft und transportiert. Hydrophobe organische Moleküle können direkt in die Blasen übergehen und mit aufsteigen.

Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Die Vesikelbildung
Mayer et al. [2, 12] konnten in Hochdruckexperimenten die Bildung von Vesikeln und eine chemische Evolution von Peptiden nachweisen. Hierbei wurden die Verhältnisse der oberen Kruste mit zahlreichen Kavitäten und einem Kaltwasser-Geysirsystem simuliert. Der Fokus der Experimente lag auf der Tiefe von ca. 1000 m, dem Übergangsbereich von ükCO2 zu Gas. Auf den Störungsflächen treten entlang von Abrisskanten eine Vielzahl von Kavitäten auf, in denen sich das Gas staut. Hierdurch entstehen Reaktionskammern mit zwei Phasen, im unteren Teil Wasser und im oberen ükCO2 oder gCO2. Liegt ükCO2 vor, führt die Druckerniedrigung während eines Geysirausbruchs in der Grenzregion zum Phasenwechsel. Es entsteht CO2 Gas, in dem gelöstes Wasser zu einem Nebel kondensiert (entspricht salzfreiem Kondenswasser). Gleiche Prozesse konnten mit Stickstoff als Gas unter milderen Bedingungen simuliert werden.
V1 Video mit Blick in die Hochdruckzelle aus Abb. 1: Untere Hälfte Wasser, das mit einem Magnetrührer gerührt wird. Obere Hälfte ükCO2 /gCO2
Der über dem Wert für überkritisches CO2 liegende Druck wird langsam reduziert. Mit Unterschreitung des überkritischen Zustandes bilden sich Nebeltröpfchen aus dem geringen Anteil des im ükCO2 gelösten Wassers. Sie bekommen im Gas eine erste Hülle aus Lipiden. Durch Absinken auf die Wasseroberfläche, die auch mit Lipiden besetzt ist, bildet sich eine zweite Hülle. Im Ergebnis liegen Vesikel vor, die aus den kondensierten Wassertröpchen und einer Lipidmembran bestehen.
Die Nebel Tröpfchen werden im Gas mit Lipiden eingehüllt, deren hydrophiler Kopf zum Wasser ausgerichtet ist, während der hydrophobe Schwanz zum Gas zeigt. Durch Absinken auf die Wasseroberfläche, die auch mit gleich orientierten Lipiden besetzt ist, bildet sich schlagartig eine zweite Hülle. Wie die Experimente gezeigt haben ist dies der Startpunkt der Vesikelbildung, die aus den kondensierten Wasser Tröpfchen und einer Lipidmembran bestehen. Hinzugegebene Aminosäuren, die im hydrothermalen Umfeld gebildet werden können, haben sich während der Druckabnahme zu Peptiden verknüpft und wechselwirkten mit der Membran der gebildeten Vesikel. Die Verknüpfung erfolgte spontan, ohne vorherige Aktivierung der Aminosäuren und ohne Hilfestellung von Katalysatoren. Allein die hohe Zunahme der Entropie ist hierbei die steuernde Größe. Die längste Kette, die von der AG Prof. Oliver J. Schmitz bestimmt wurde bestand aus 18 Einheiten.
V2 MD-Simulation als Beispiel für das Verhalten von hydrophoben Peptiden (hier 8 Aminosäuren) in einer Membran. Die blau gepunkteten Bereiche sind Wassermoleküle. Die Membran besteht aus Lipiden mit grauen und grünen hydrophilen Köpfen und grünen, hydrophoben Schwänzen, die nach innen gerichtet sind. Clip mit freundlicher Genehmigung von Maria Davila Garvin und Christian Mayer, Physikalische Chemie der Universität Duisburg-Essen. Fragen und Hinweise direkt an sie: maria.davila@uni-due.de christian.mayer@uni-due.de
Mit einer erneuten Druckerhöhung durch zurückfließendes Wasser nach Ende des Ausbruchs werden die meisten Vesikel wieder zerstört. Am Ende konnten Mayer et al. zeigen, dass eine chemische Evolution durch zyklische Wiederholung der Druckschwankungen möglich ist, die zur gegenseitigen Stabilisierung von Peptiden und Vesikeln führt. Die unendlich erscheinende Vielzahl der Kombinationsmöglichkeiten bei der Peptidbildung lässt aber keine Wiederholung gleicher Sequenzen in den Aminosäureketten erwarten. Bei einer Kettenlänge von 8 Aminosäuren aus den 12, die hydrothermal gebildet werden können, ergibt sich eine Variation von ca. 430 Mio. Möglichkeiten. Was fehlt, ist eine Speicherung der Information über die Abfolge, so wie sie in der später gebildeten RNA bzw. DNA zu finden ist.
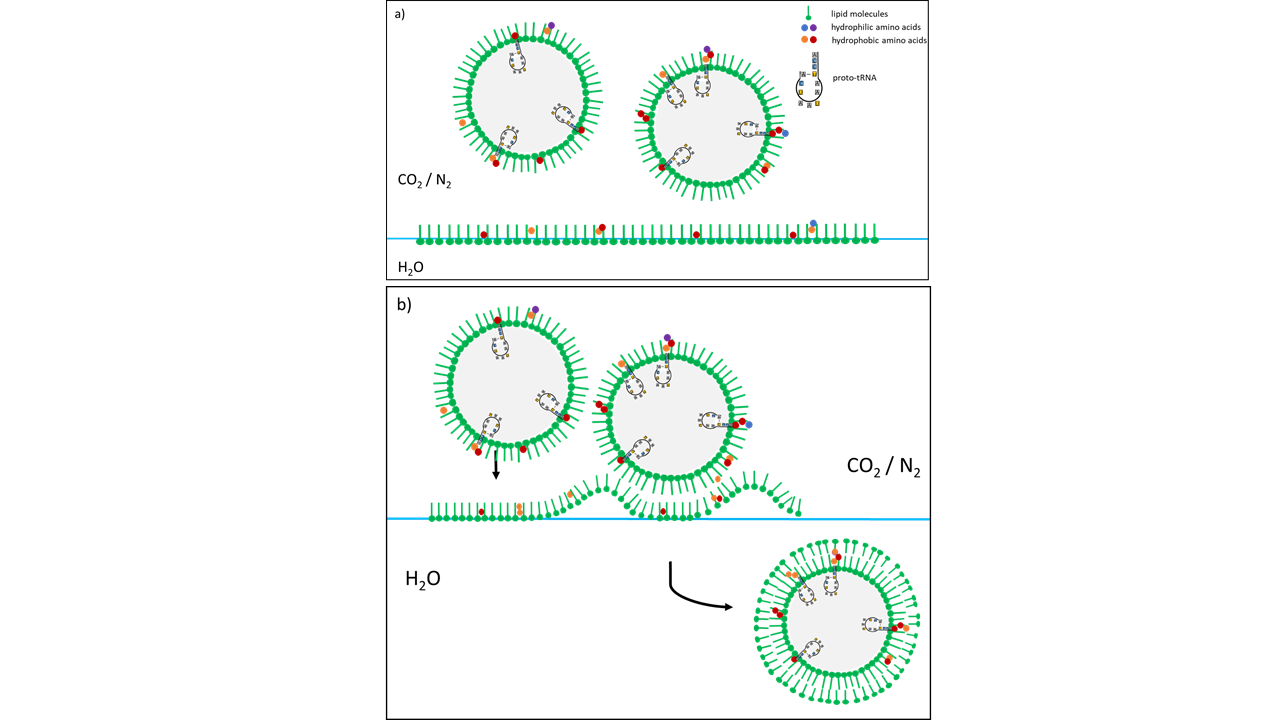
Abb. 10 Vesikelbildung nach Druckreduktion infolge eines Kaltwasser-Geysir Ausbruchs. a) Nebeltröpfchen werden mit einer Hülle aus Lipiden ummantelt. b) Im Kontakt mit Lipiden auf der Wasseroberfläche kommt es zu einer zweiten Umhüllung (auf Grundlage von Mayer et al. 2015 [2]). Die Vesikel bestehen aus destilliertem Wasser mit einem Anteil organischer Moleküle. Der Unterschied in der Salzkonzentrationen außen (hoch) und innen (salzfrei) führt zu einer Instabilität, die zu einer schnellen Zerstörung während des nächsten Geysirausbruchs führt. Der gleiche Vesikelbildungsprozess verläuft mit Wasserspritzern, die bei einem turbulenten Geysirausbruch über die Wasseroberfläche geschleudert werden. Die sich bildenden Vesikel sind deutlich größer, besitzen eine wesentlich höhere Konzentration organischer Moleküle, aber die gleichen Salzgehalte, wie das Umgebungswasser. Hierdurch sind sie stabiler als die Vesikel aus den Nebeltröpfchen und haben höhere Konzentrationen organischer Moleküle für Reaktionen. Dargestellt sind Aminosäuren (farbige Punkte) und eine spezielle RNA (s.o.), die mit der Membran wechselwirken.
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Die Bedeutung der Vesikelbildung zeigt sich in gleich mehreren Eigenschaften, die in der Entwicklung des Lebens eine bedeutende Rolle gespielt haben.
- Einerseits wird deutlich, wie leicht Vesikel als Vorläufer der Zellkompartimente gebildet werden können.
- Andererseits tritt eine Wechselwirkung zwischen Membran und Molekülen ein, die zu speziellen Reaktionen und einer chemischen Evolution führen können.
- Weiterhin ist die Art der Vesikelbildung geeignet, große Molekülkomplexe sowie Molekülgemische, die im freien Umfeld der Kavitäten gebildet werden, während der Tröpfchenbildung aufzunehmen.
Es ergeben sich aus den bishier vorgestellten Ergebnissen weitreichende Überlegungen für neue Fragestellungen und experimentelle Designs. So lassen sich neben den bisher verwendeten kanonischen L-Aminosäuren alternativ D-Aminosäuren für die gleiche Versuchsreihe einsetzen. Es ist hierbei interessant, ob sich durch die chemische Selektion in den Peptiden mit D-Aminosäuren ähnliche Sequenzen mit einer chemischen Evolution ergeben, wie mit L-Aminosäuren. Eine weitere Versuchsreihe kann mit racemischen Gemischen durchgeführt werden, bei denen die Verhältnisse der beiden Versionen gleich sind.
Von besonderem Interesse sind die Sequenzen der D-Peptide für die pharmazeutische Forschung, in der es um die Entwicklung neuer Antibiotika geht. Hierfür müssen zusätzliche Komponenten für spezielle selektive Schritte in den Vesikelbildungsprozess integriert werden.
Möglicherweise kann ein Versuchsdesign mit D-Aminosäuren und L-Ribose einer RNA ein Anfang für die Entwicklung einer Spiegelbild Zelle sein (s. auch: Confronting risks of mirror life, DOI: 10.1126/science.ads9158).
Das hypothetische Modell zur Speicherung von Information
Für die Speicherung der Information über die Aminosäuresequenzen in den Peptiden ist ein vermittelndes Molekül erforderlich gewesen, das einer einfachen Version einer tRNA entsprochen haben muss (s.o). Diese Proto-tRNA wird in dem hier vorgestellten Modell für die Entwicklung der ersten Zelle vorausgesetzt. Die Nutzung der DNA als Träger der Erbinformation wird für in einen späteren Zeitpunkt angenommen.
Der Beginn der Informationsspeicherung
Die Bildung von Vesikeln unter Druck- und Temperaturbedingungen, wie sie in ca. 1000 m Tiefe einer kontinentalen Kruste der jungen Erde entsprechen (s.o.), ist im Labor mehrfach nachgewiesen worden [2, 4]. Bis zu 12 Aminosäuren können unter hydrothermalen Bedingungen in der Kruste entstehen [15, 16], wobei die Häufigkeiten in Abhängigkeit der Bildungsbedingungen unterschiedlich sind. Die (hydrothermal bildbaren) hydrophoben Aminosäuren Isoleucin, Valin, Leucin und Phenylalanin könnten bei der Vesikelbildung theoretisch in die Membran aufgenommen werden, wobei es aber energetische Hürden gibt. Dies ist nicht der Fall, wenn sich mehrere gleiche oder ähnliche Aminosäuren zu kurzen Peptiden verbinden. In dieser Form ist eine Aufnahme in die Membran leicht möglich. Die hydrophilen Aminosäuren Threonin, Serin, Prolin, Glutaminsäure, Asparaginsäure und Lysin verbleiben im Wasser der Tröpfchen, sofern sie nicht in Peptide mit überwiegend hydrophoben Aminosäuren eingebunden sind. Glycin, als einzige nicht chirale Aminosäure, ist ein Sonderfall und kann unterschiedliche Positionen einnehmen.
Die Vesikel bestehen aus destilliertem Wasser mit einem Anteil organischer Moleküle. Der Unterschied der Salzkonzentrationen innen und außen führt zu einer Instabilität, die zu einer schnellen Zerstörung während des nächsten Geysirausbruchs führt. Der gleiche Vesikelbildungsprozess verläuft mit Wasserspritzern, die bei einem turbulenten Geysirausbruch in der Tiefe über die Wasseroberfläche der Kavität geschleudert werden. Die sich hierbei bildenden Vesikel sind deutlich größer, besitzen eine wesentlich höhere Konzentration organischer Moleküle, aber die gleichen Salzgehalte, wie das Umgebungswasser. Hierdurch sind sie stabiler als die Vesikel aus den Nebeltröpfchen und haben höhere Konzentrationen organischer Moleküle für Reaktionen. Die in diesen Vesikeln gebildeten Reaktionsprodukte unterscheiden sich zu denen der kleineren Vesikel mit destilliertem Wasser in Qualität und Quantität.
Hydrophobizität - eine steuernde Größe
An der oben skizzierten einfachen Proto-tRNA (Abb. 8) ist mit der
Basenfolge
CCA die Base Adenin genau an der Spitze des hier als ungepaart vorliegenden Strangs positioniert. Der Akzeptorarm kann durch die Hydrophobizität des Adenins in die Membran eindringen und gelangt so in Kontakt zu den hydrophoben Aminosäuren (Abb. 11). Eine Verknüpfung an der 2‘-OH Position der endständigen Ribose im Rahmen des Geysirzyklus ist jetzt möglich.

Abb. 11 Unterschiedliche Eindringtiefen des Akzeptorarms in die Membran in Abhängigkeit der Hydrophobizität des Anticodons
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
In der chemischen Evolution, die zum „ACC“ Arm geführt hat, müssen zahlreiche Variationen ausgebildet worden sein, die aufgrund von Molekülgröße und/oder Grad der Hydrophobizität nicht oder weniger geeignet waren (Abb. 13):
Die Frage ist, wie eine spezifische Verbindung der entsprechenden Aminosäure in Bezug zum Basentriplett des Anticodons erreicht werden konnte. Am Basentriplett, das letztlich den Code vorgibt, liegen immer drei von vier möglichen Basen vor, die sich hinsichtlich ihrer Hydrophobizität unterscheiden [10]. Je nach Art der beteiligten Basen und ihren Positionen auf dem Triplett ergeben sich unterschiedliche Werte für die entropische Kraft, die im Zuge des hydrophoben Effekts zu einem unterschiedlich tiefen Eindringen des Akzeptorarms in die Membran beiträgt (https://de.wikipedia.org/wiki/Hydrophobie).
Das bedeutet, je stärker die Hydrophobizität im Bereich des Anti-Codons ist, desto weiter ragt die Spitze der Proto-tRNA in die Membran. Kommt sie dort mit einer endständigen Aminosäure eines Peptids in Kontakt, kann die Aminosäure bei gleichzeitiger Abspaltung an die Spitze der Proto-tRNA gebunden werden. Durch die Abtrennung der Aminosäure von der Kette wird die Entropie erhöht und somit die Reaktion begünstigt. Die Verknüpfung bindet die Aminosäure an der 2‘-OH Position. Mit entsprechenden Kombinationen der Basen am Anticodon werden unterschiedliche, sehr fein abgestimmte Positionen des CCA-Arms in der Membran erreicht, wodurch unterschiedlich hydrophobe Aminosäuren mit unterschiedlichen Positionen in der Membran (während eines Druckabfalls mit Phasenwechsels des Gases) verknüpft werden. Die Aufenthaltswahrscheinlichkeit für die am stärksten hydrohopen Aminosäuren ist in der innersten Zone im Mittel am größten. Liegen hydrophilere Basen am Anticodon vor, reicht der CCA Arm nicht so weit in die Membran, sodass die hydrophilen Aminosäuren aus dem Wasserbereich an der 3‘-OH Position verknüpft werden. In einer mittleren Position können keine Aminosäuren angebunden werden. Die entsprechende Kombination entspricht den heutigen Stop Positionen beim Ablesevorgang der mRNA im Ribosom.
Unterstützung
Das Modell schien gewagt, gab es doch neben der vermuteten Ursache des entropischen Effekts keine Belege, dass eine RNA, wenn auch nur mit einem kurzen einsträngigen Abschnitt, in eine Vesikelmembran eintauchen kann. Die gängige Meinung war, dass sich eine RNA maximal parallel hierzu anordnen könne. Mitte 2025 erschien von Czerniak und Saenz ein Review, dass die bisherigen Arbeiten zu Reaktionen von RNA mit Lipiden zusammenfasst und bewertet [26]. Die Autoren stellen fest, dass trotz des stark hydrophilen Charakters der Nukleinsäuremoleküle die Existenz von Nuklebasen in unmittelbarer Nähe des hydrophoben Kernbereichs von Lipidmembranen beobachtet wurden und dass Guanin, Adenin und Uracil mit den Lipidketten interagieren können. Sie schließen aus den Beobachtungen der zitierten Bearbeiter, dass entfaltete einzelsträngige RNA freiliegende Nukleinsäurebasen aufweisen, die mit den tieferen Regionen von Phospholipidmembranen reagieren können [hierzu s. . Michanek et al. (2012); 27]. Ein solcher Mechanismus sei bei doppelsträngigen Spezies, bei denen die Nukleobasen in RNA-Helices eingebettet sind, nicht möglich. Bemerkenswert sei darüber hinaus, dass in einer Reihe von Experimenten eine Anzahl von RNAs gefunden wurden, die spezifisch an hydrophobe Aminosäuren wie Valin, Tryptophan, Phenylalanin und Isoleucin binden. Dies bestätigt, so ihr Fazit, dass es grundsätzlich möglich ist, dass RNA-Spezies an hydrophobe Ketten von Lipiden binden können.
Aus mechanischer Sicht ist das Fixieren eines Fingers in einem plastischen Medium günstiger, wenn die Spitze verdickt ist (vergleichbar mit einem Lolli, der in einen festen Pudding gesteckt wird). Mit der Basenkombination „ACC“ am Akzeptorarm folgen auf Adenin, das zwei Ringstrukturen hat (Purin-Grundgerüst), mit Cytosin zwei kleinere Basen mit nur einer Ringstruktur (Pyrimidin-Grundgerüst). Diese Kombination hat sich vermutlich als die Günstigste erwiesen (Abb. 12).
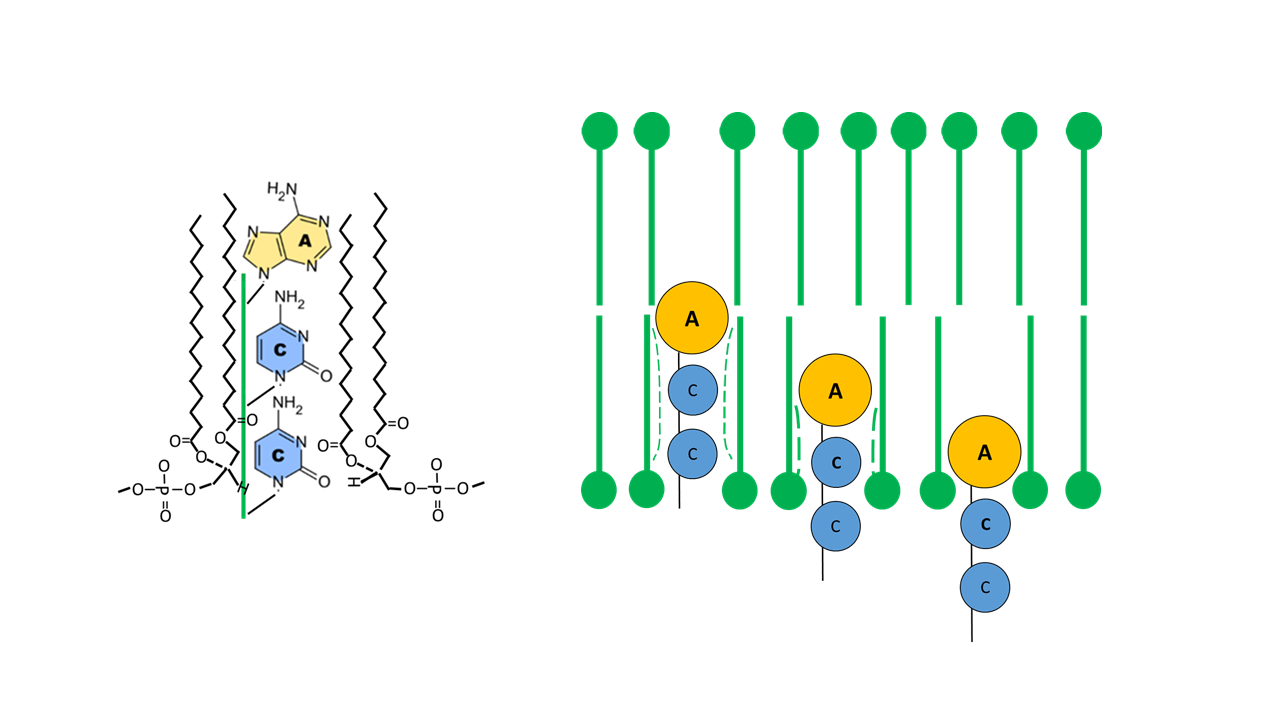
Abb. 12 Fixierung des ACC Arms in der Membran (Lolli-Struktur zur optimalen Einbindung, Funktion der Membran als Katalysator)
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Der ACC oder CCA Arm
In der chemischen Evolution, die zum „ACC“ Arm geführt hat, müssen zahlreiche Variationen ausgebildet worden sein, die aufgrund von Molekülgröße und/oder Grad der Hydrophobizität nicht oder weniger geeignet waren (Abb. 13):
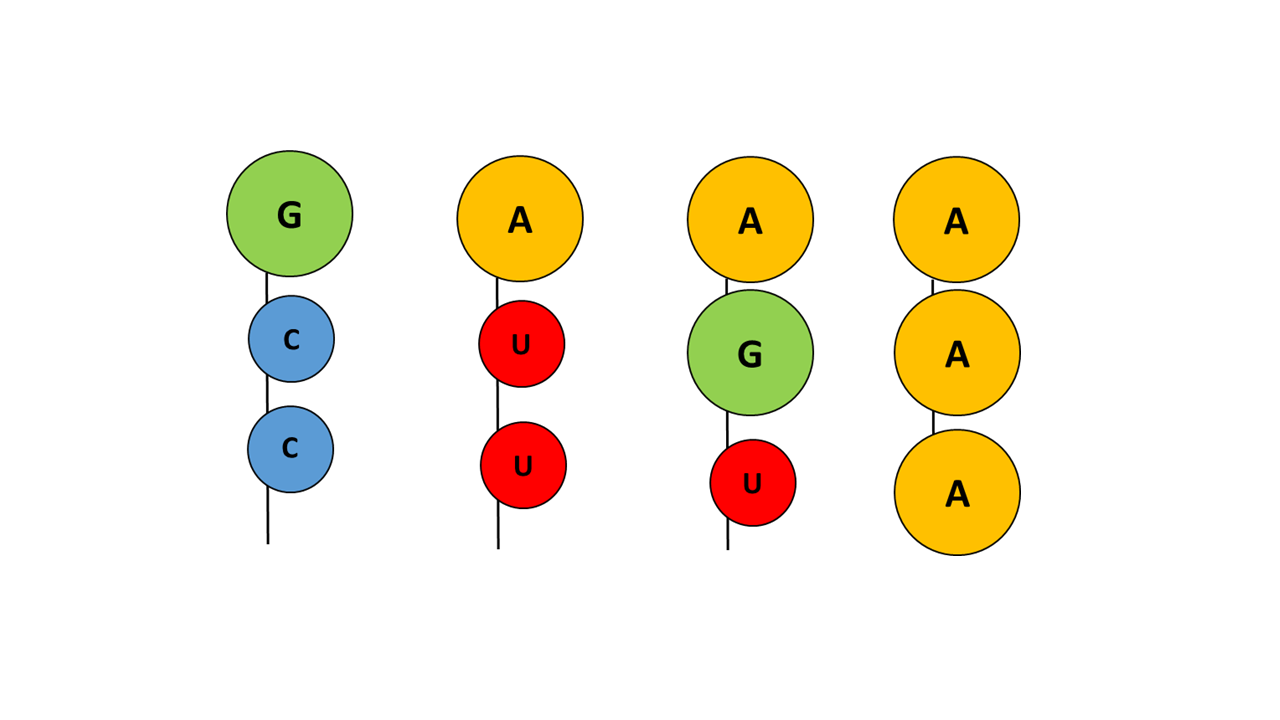
Fig. 13 Variationen verschiedener Basenkombinationen des Akzeptorarms, die sich nicht durchgesetzt haben
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Mit Guanin an der Spitze liegt eine hydrophile Base vor, die ein Eintauchen in die Membran blockiert, unabhängig davon, welche Nukleinbasen folgen. Mit Adenin an der Spitze und der kleineren Pyrimidin Base Uracil ist ein Eindringen des Akzeptorarms in die Membran möglich, allerdings ist Uracil stärker hydrophil als Cytosin, sodass die Kombination mit Cytosin günstiger ist. Liegt nicht nur an der Spitze sondern auch an weiteren Positionen die Nukleinbase Adenin vor, ist die Fixierung in der Membran zu stark, sodass eine Feinsteuerung der Eindringtiefe durch das Anticodon nicht mehr gegeben ist.
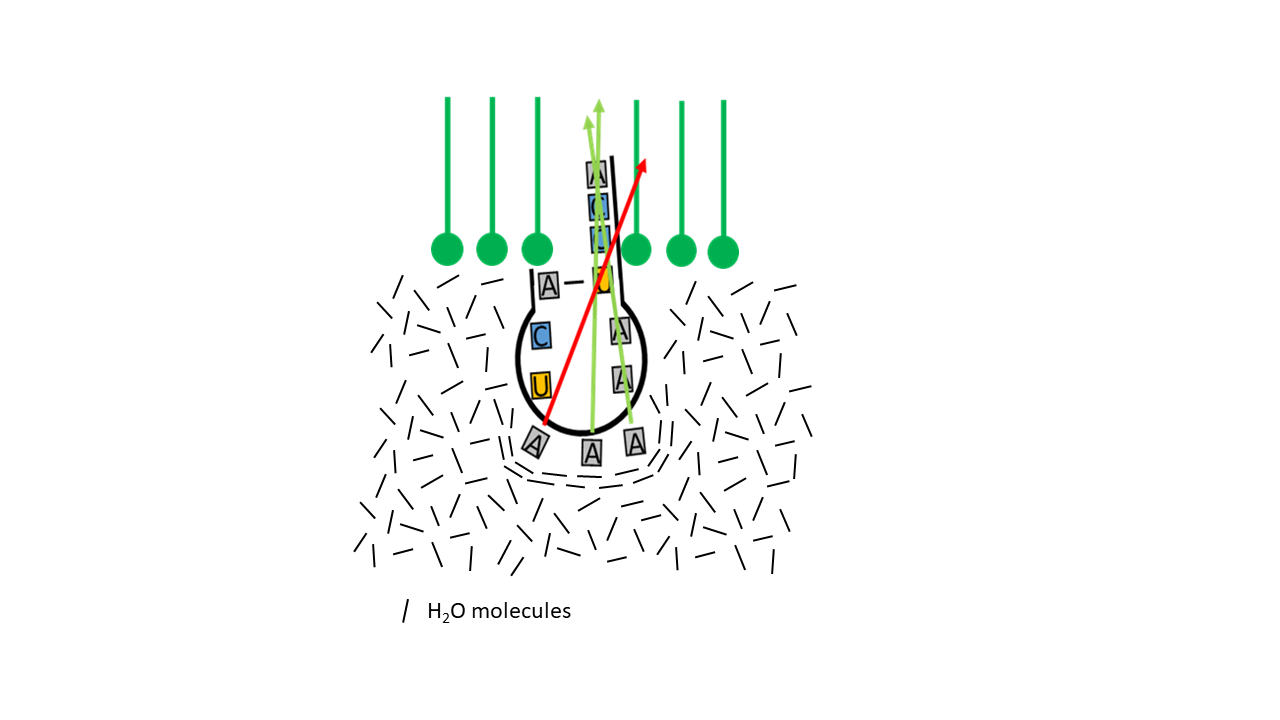
Abb. 14 Richtung der Kraftvektoren des hydrophoben/entropischen Effekts in Abhängigkeit der Molekülstruktur
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Wobble Position
Eine Besonderheit für die Spezifität des genetischen Codes zeigt sich in der Variabilität der dritten Base des Codons der mRNA (5‘ -> 3‘) bzw. der des Anticodons (3‘ -> 5‘). Bei Tripletts, die für dieselbe Aminosäure codieren, gibt es häufig nur einen Unterschied in der Variation der dritten Base (--> Wobble-Paarung). Die Bedeutung für die Spezifität der dritten Base ist folglich geringer als für die erste und zweite Base.
Zu überlegen ist, ob es einen Zusammenhang zum hydrophoben/entropischen Effekt gegeben haben kann. Werden zum Beispiel die wirkenden Kräfte an einer an die Membran gekoppelten Proto-tRNA anhand von Kraftvektoren betrachtet (Abb. 14), ist, durch die Molekülkonfiguration bedingt, der Einfluss der ersten beiden Basen auf die Eindringrichtung des Akzeptorarms größer, als der der dritten Base.
Für Hinweise, Korrekturen oder Diskussionen: cogita.ama@gmail.com
Eine erste Idee - aber unrealistisch
Mit diesen Zusammenhängen ließe sich ein Prozess ableiten, der durch eine enge Besetzung von verschiedenen Proto-tRNAs an einer Innenseite einer Vesikelmembran eine Speicherung einer Aminosäuresequenz in einer RNA zulässt (Abb. 15). Hierbei würde die Krümmung der Membran eng benachbarte Positionen der Anticodons ermöglichen, sodass eine komplementäre Anlagerung von Codons zur Bildung einer RNA (Proto-mRNA) führen könnte. Ein Problem bereitet hier allerdings die Händigkeit der Aminosäuren bzw. der Ribose, für die eine Selektion für ein homochirales Peptid fehlt. Bei längeren Ketten ergeben sich unüberschaubar viele Kombinationsmöglichkeiten, die entsprechende Zeiträume zum Ausprobieren erfordert. Ein effektiverer Vorgang wird dann erzielt, wenn alternativ die Proto-tRNA an der einzigen Doppelbindung getrennt wird. Sie liegt hiernach als Einzelstrang vor und kann die Funktion einer Proto-mRNA übernehmen (s.u.).
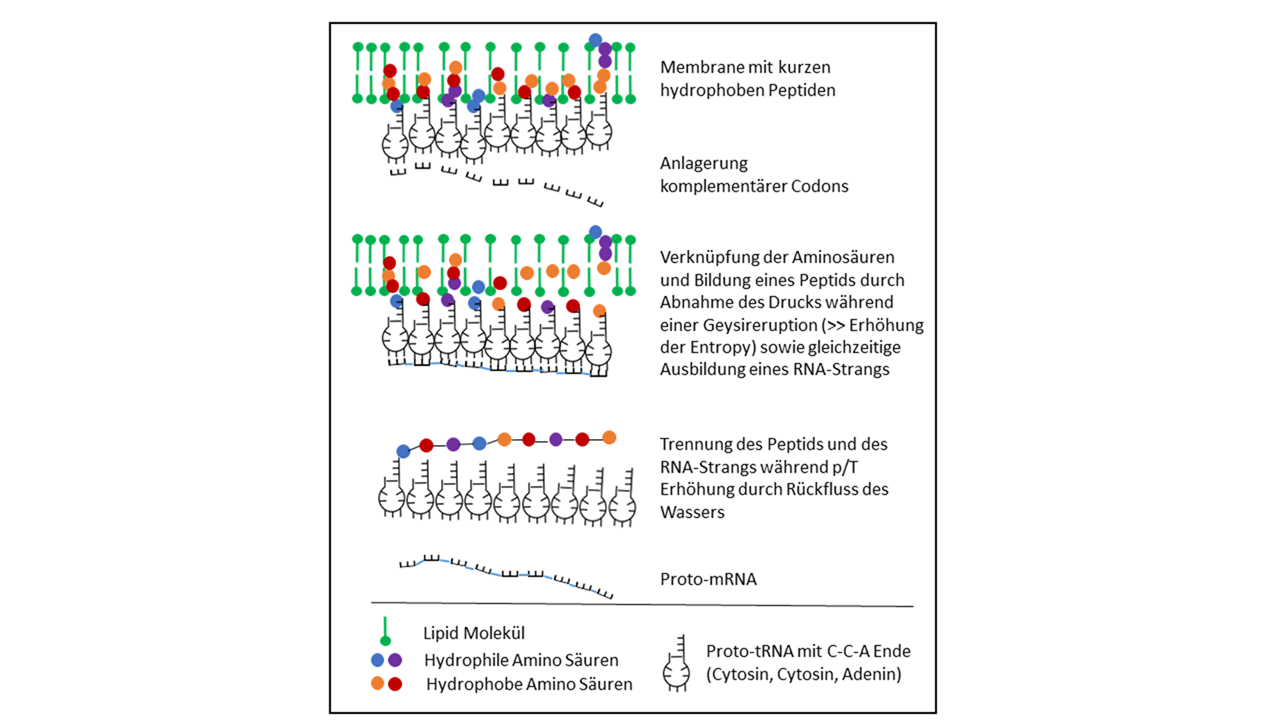
Abb. 15 Eintauchen der Proto-tRNA in die Vesikelmembran mit dem Akzeptorarm CCA an der Spitze in Abhängigkeit von der Hydrophobizität, die vom Anti-Codon vorgegeben wird. Nach Verknüpfung der Aminosäuren mit den Proto-tRNAs (hydrophobe AS gelb und rot sind an kurze Peptide in der Membran gebunden) trennen sich die Aminosäuren von den kurzen Peptiden und verknüpfen sich untereinander zu einem längeren Peptid. Die eng zusammenliegenden Anti-Codons bilden den Informationsspeicher für die Sequenz des Peptids und dienen als Template für eine Proto-mRNA, die durch Anlagerung komplementärer Nukleotide entsteht und sich anschließend abtrennt. – Dieses Möglichkeit wurde nicht weiter betrachtet, da die Variationsmöglichkeiten mit immer gleichzeitig auftretenden beiden Händigkeiten der Ribose ins Unendliche geht.
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
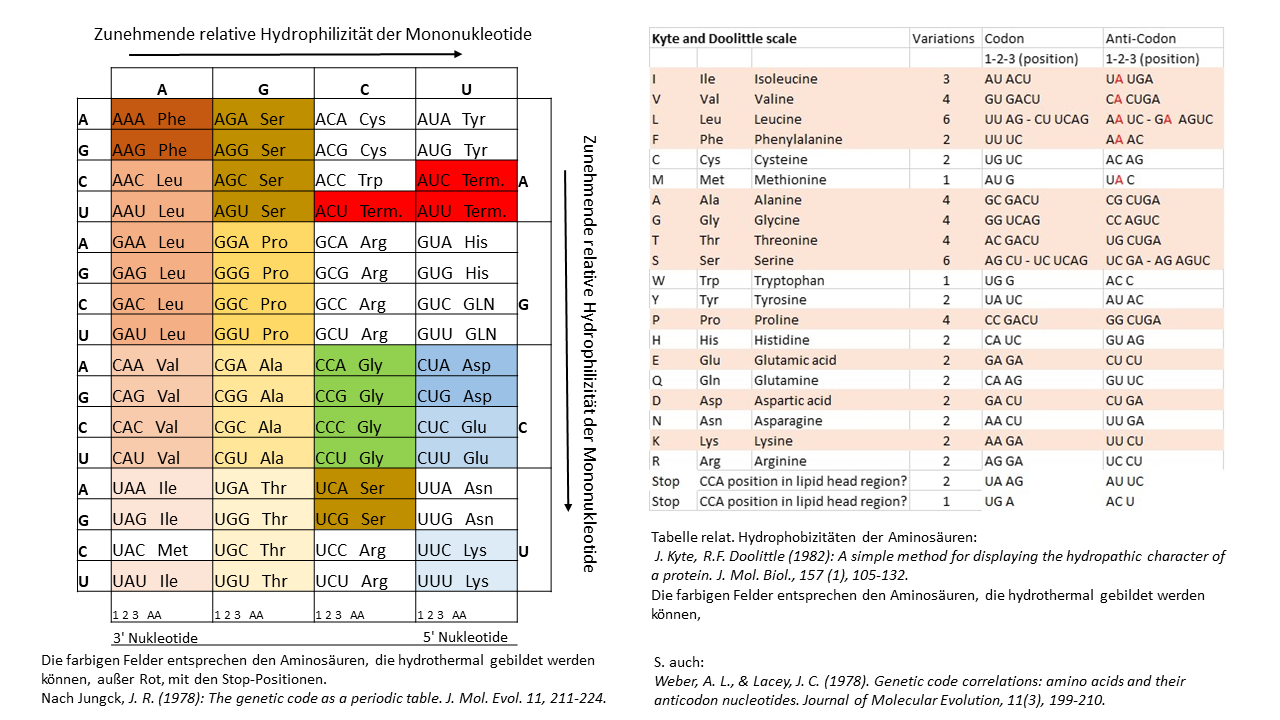
Das vorgestellte Modell der Positionierung der einsträngigen Proto-tRNA Spitze in einer Membran erfordert eine strenge Korrelation in den Hydrophobizitäten der Aminosäuren mit den Nukleobasen bzw. der Anticodons. In der Tab. 1 wird beim Vergleich der Anticodon Belegungen mit den zugehörigen Aminosäuren deutlich, dass alle hydrothermal bildbaren hydrophoben Aminosäuren ein Adenin in der Mittelposition bzw. weitere im Randbereich besitzen (rechts). Die hydrophilen Aminosäuren haben eine entsprechend entgegengesetzte Belegung. Bereits Weber and Lacey [17] entdeckten 1978 diese auffällige Korrelation, ohne jedoch eine Erklärung hierfür anbieten zu können. Besonders deutlich wird dieses Verhältnis in der Tabelle von Jungck 1978, wenn nur die hydrothermalen Aminosäuren (farbige Felder bis auf Codons der Stop Positionen) berücksichtigt werden.
Tab. 1 linke Seite nach Jungck, J. R. (1978)[22]: In dieser Tab. werden die Verhältnisse besonders deutlich. Dargestellt sind die Verhältnisse von drei der vier Basen des jeweiligen Anticodons zur codierten Aminosäure in Abhängigkeit der Hydrophilizität (1. Base links ist jeweils die Wobble Position). Farblich hervorgehoben sind die Aminosäuren, die in hydrothermalen Systemen gebildet werden können (Ausnahme Rot für Stop-Positionen). Die hydrophilen Aminosäuren (Blautöne) liegen in der Ecke mit den am stärksten hydrophilen Anticodons. Gegenübergestellt zum Vergleich Tab. 1 (rechte Seite). Eine ausreichende Erklärung für diese eindeutigen Zusammenhänge gab es bisher nicht.
Tab. 1 rechte Seite: Die kanonischen Aminosäuren mit abnehmender Hydrophobizität nach Kyte und Doolittle [20]. Hydrothermal bildbare Aminosäuren sind farblich hinterlegt. Bei den hydrophoben (hydrothermalen) Aminosäuren ist die zweite Position des Anti-Codons an der tRNA (unterstes Ende) jeweils mit dem hydrophoben Adenin (in Rot) besetzt. Die tRNAs der hydrophilen AS besitzen in dieser Position die hydrophilen Basen G, U oder C.
Die unterschiedlichen Proto-tRNAs konnten relativ spezifisch anhand der Hydrophobizität des Anticodons mit Aminosäuren verknüpft werden. Gleichzeitig ließ sich leicht die einzige Doppelbindung der tRNAs trennen, sodass ein Einzelstrang existierte. Hierbei ist die Paarung Adenin-Uracil (A-U) weniger stark gebunden, als die Bindung Guanin-Cytosin (G-C), die eine Wasserstoffbrückenbindung mehr besitzt. Der Einzelstrang stellte einen Informationsstrang dar, der zu Beginn durch seine zufällige Nukleotidabfolge keine Bedeutung hatte. Da zur gleichen Zeit beladene tRNAs vorgelegen haben, die mit der G-C Basenpaarung bei einer Temperaturerhöhung länger stabil waren als die Proto-tRNAs mit der Bindung A-U, konnten diese den `A-U´ Strang als Proto-mRNA nutzen und ein Viererpeptid bilden, sofern passende Verhältnisse vorlagen.
Einige der "tRNA-Templates" konnten nicht für ein Peptid genutzt werden, da sie Plätze für Aminosäuren zur Verfügung stellten, die noch nicht existierten. Dies gilt auch für die komplementären Stränge, die bei der Replikation (die als Voraussetzung für das Überleben der Proto-tRNAs gilt) entstehen (Abb. 16). Der CCA Abschnitt steht nach der Replikation im komplementären Strang zum Beispiel für die Aminosäure Tryptophan, das vermutlich nicht hydrothermal gebildet wurde und erst später den Code UGG eingenommen hat. Aus Analogiegründen könnte UGG ursprünglich für Threonin gestanden haben.
Da die Proto-tRNA homochiral (sowohl als D und als L Version) vorliegen musste, um als solche gebildet zu werden und es eine bevorzugte Verknüpfung von L Aminosäuren mit D RNAs (und umgekehrt) gibt [23], waren die gebildeten Peptide auch homochiral (in gleichen Mengen mit beiden Händigkeiten). Da maximal 12 verschiede Aminosäuren mit unterschiedlichen Proto-tRNAs vorgelegen haben konnten, von denen es zum Teil mehrere für eine Aminosäure gab, standen eine Vielzahl von unterschiedlichen Templates für Viererpeptide zur Verfügung. Die Verknüpfung von zwei Viererpeptiden mit gleicher Händigkeit ergab Längen von Membranstärke. Hiermit konnten bereits Poren gebildet werden, die für einen Konzentrationsausgleich erforderlich sind. Da die Sequenzen der Peptide bereits gespeichert waren, kam es nur noch auf die Kombination der Viererketten bzw. der entsprechenden komplementären RNA-Stränge an, um erste Funktionsmoleküle zu erhalten, die katalytisch wirksam waren. Trugen diese dazu bei, weitere Funktions- und andere "nützliche" Molküle schneller zu bilden, gewannen sie den Wettlauf um Ressourcen und setzten ihre Händigkeit durch.
Betraf dieser Prozess zum ersten Mal ein Molekül, das katalytische Funktion besaß und das im weiteren Verlauf der Entwicklung z.B. als Kern komplexerer Peptide erhalten blieb, kann dies als Start des Lebens angesehen werden.
Vorteil der Nutzung der geöffneten Proto-tRNA als Template:
Werden sowohl die Einzelstränge der tRNA (sofern die Codes zu den hydrothermalen Aminosäuren passen) als auch gleichzeitig die komplementären Stränge als Templates benutzt (Abb. 16), können hieraus katalytisch wirksame Peptide gebildet werden, die in einer späteren DNA eine bidirektionale genetische Kodierung (Sense/Antisense-Ausrichtung) aufweisen.
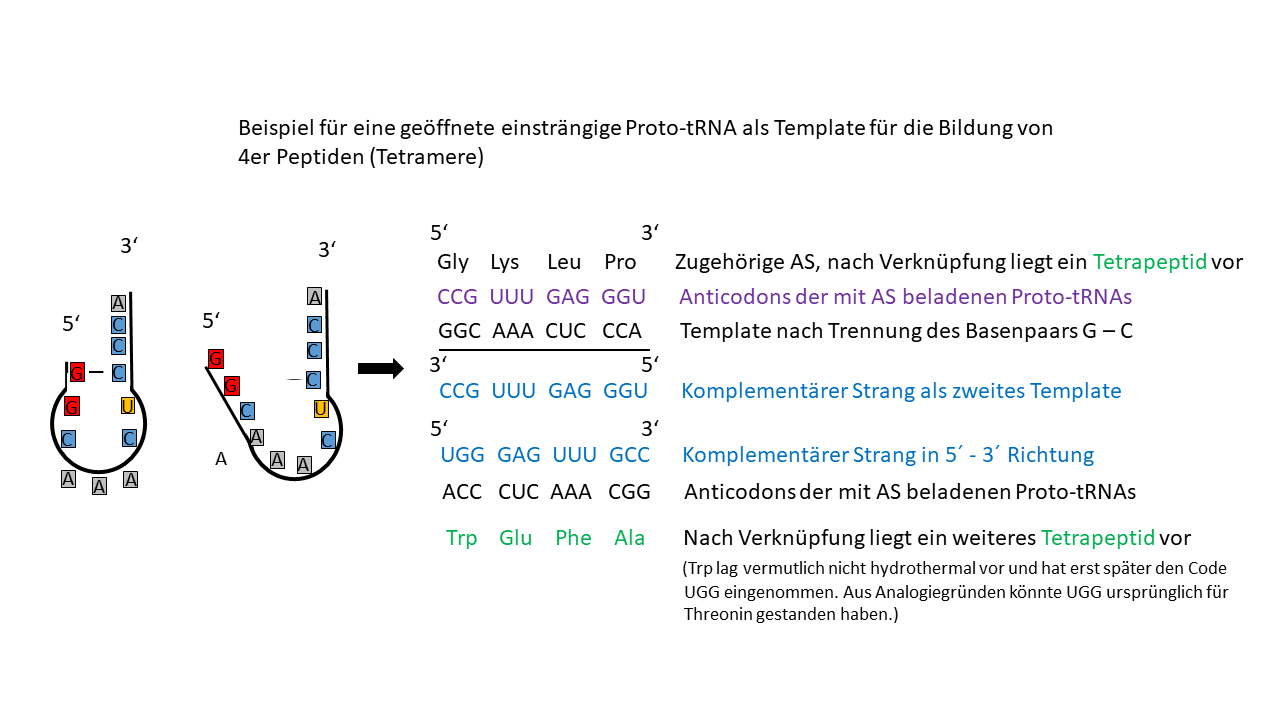
Abb. 16 Proto-tRNA als Einzelstrang, der als Template fungieren kann. Ein weiteres Template ergibt sich aus dem komplementären Strang. Von beiden können Tetramere gebildet werden, die durch die tRNA homochiral sind. Allerdings war die Einzelstrangbildung mit dem Basenpaar A-U eher bei Temperaturerhöhung zu erreichen, sodass beladene tRNAs mit G-C Paarung diese bereits als Template nutzen konnten.
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
Wie oben aufgeführt gibt es heute zwei Klassen von Synthetasen, die die spezifische Beladung der tRNAs vornehmen. Die zur Klasse-I-Synthetase gehörenden Aminosäuren, die am 2´- OH Ende der endständigen Ribose verknüpft werden, sind im Allgemeinen größer und weniger polar (z.B. die hydrophoben AS), die der Klasse-II-Synthetase, die am 3´- OH Ende angebunden werden, sind kleiner und polarer (z.B. die hydrophilen AS). Biochemische, bioinformatische und proteintechnische Experimente stützen die Hypothese, dass die beiden Klassen von entgegengesetzten Strängen desselben Vorfahrengens abstammen [25]. Dies würde die Überlegung stützen, dass die Grundlage der Entwicklung der heutigen Aminoacyl-tRNA Synthetasen auf eine sehr frühe Phase zurückgeht, die mit der oben geschilderten Template Bildung zusammenhängt.
Tab. 2 Komplementäre Gruppen von Aminosäuren, die einen Zusammenhang bei der Verwendung der Templates von Proto-tRNAs und deren komplementären Strängen zeigen können.
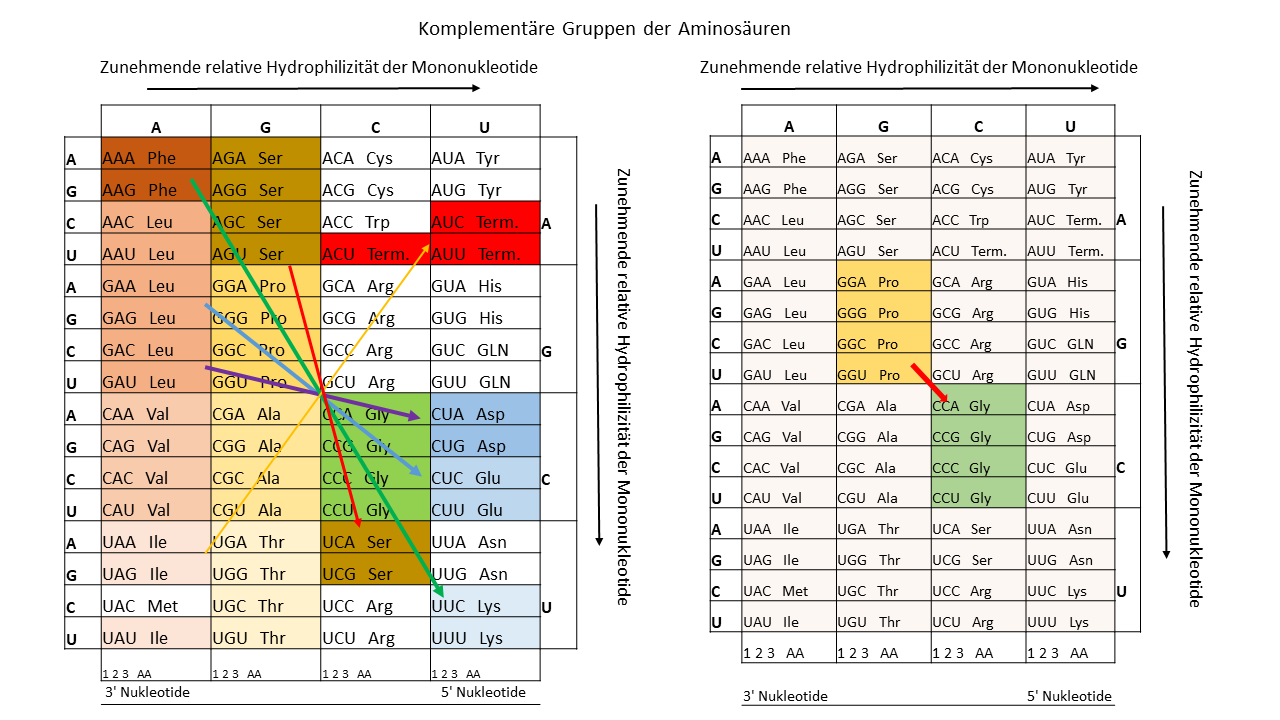
Dieses Werk ist lizenziert unter einer Creative Commons Namensnennung - Nicht-kommerziell - Weitergabe unter gleichen Bedingungen 4.0 International Lizenz.
ATP
Von besonderem Interesse ist die Bildung von ATP (Adenosintriphosphat), das in heutigen Zellen der wesentliche Energieträger ist. Es wird in einem komplexen Vorgang gebildet, der sich erst im Laufe der Evolution entwickelt hat. Für viele Reaktionen wäre zu Beginn der Lebensentstehung ATP als Energiequelle interessant. Wie oben gezeigt, kann die Vesikelmembran als Katalysator wirken. Wenn die Proto-tRNA mit Adenin an der Spitze in die Membran eindringen kann, muss dies auf für das AMP (Adenosinmonophosphat) möglich gewesen sein. Unter der Voraussetzung , dass erforderliche Moleküle vorhanden waren, lässt sich die Bildung von ATP postulieren. Entsprechende Experimente hierzu sind vorstellbar.
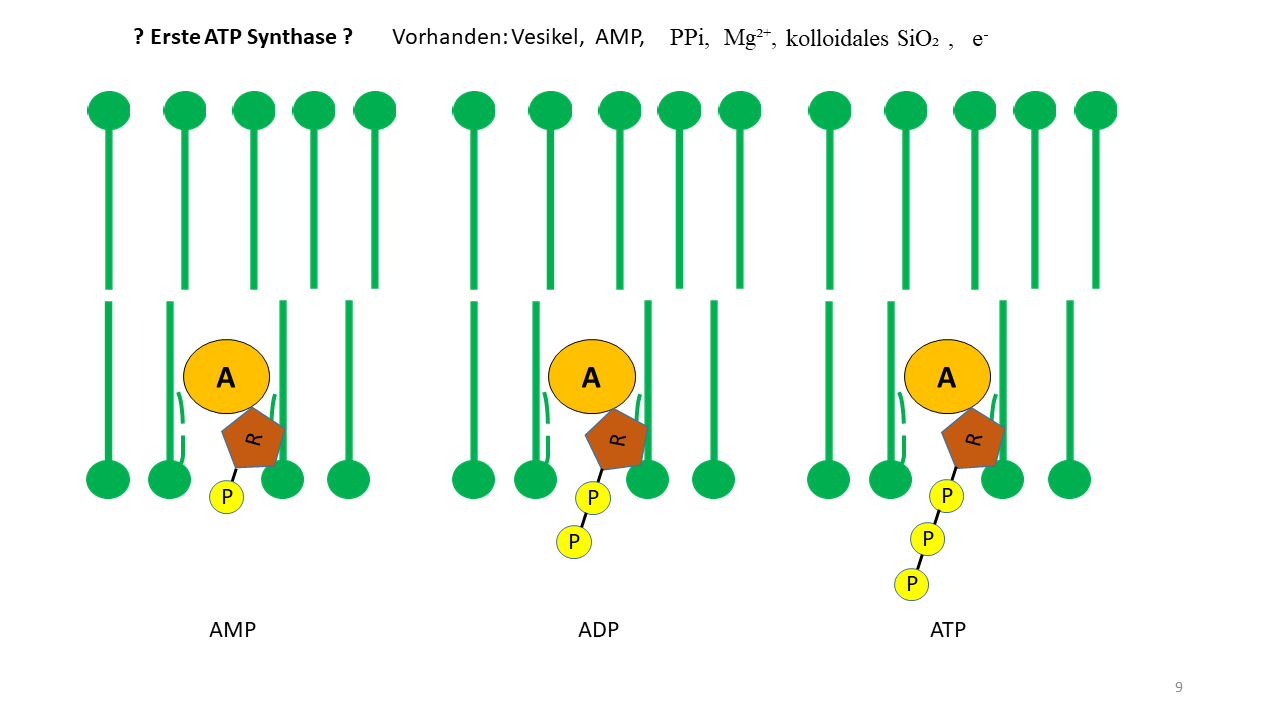
Abb. 17 Membran als Katalysator für die Positionierung von Adenosinphosphaten. Durch die die Fixierung von AMP (Adenosinmonophosphat) ist die Bildung von ADP (Adenosindiphosphat) und ATP (Adenosintriphosphat) denkbar. Vorhanden sein mussten neben AMP, Phosphat, MG²+, kolloidales SiO2 und e- durch Piezoelektrizität, was der Fall war.
Konsequenzen
Folgende Annahme kann aus den oben angeführten Zusammenhängen getroffen werden: Sowohl die Einzelstränge der Proto-tRNA aus 12 Nukleobasen als auch gleichzeitig die komplementären Stränge konnten als Templates benutzt werden, mit denen sich katalytisch wirksame Peptide bildeten.
Die 4 Tripletts jeder der möglichen Proto-tRNAs (es gab mehr als 65.000 Variationsmöglichkeiten, wobei die gleiche Anzahl an tRNAs mit der anderen Händigkeit hier nicht diskutiert sind) waren noch ohne Informationsinhalt, die für ein Funktionsmolekül notwendig waren. Es waren zufällige Kombinationen, aus denen Tetramere hervorgehen konnten. Bedingung war allerdings, dass für die jeweiligen Tripletts auch passende tRNAs und zugehörige Aminosäuren vorhanden sein mussten, da es zu Beginn noch nicht alle Aminosäuren in diesem Environment gab. Wurde diese RNA immer wieder kopiert, blieb auch die Information über die Sequenz der entsprechenden Tetramere erhalten.
Und die Chiralität?
Die Entwicklung der chemischen Evolution zu D-Ribose in der RNA und L-Aminosäuren in den Peptiden könnte möglicherweise durch einen zufälligen Überhang eines Enantiomers zu einem bestimmten Zeitpunkt erklärt werden, was aber schwer nachzuvollziehen ist. Grundsätzlich existieren immer gleiche Mengen an Molekülen in der D- und L-Version, sodass alle Reaktionsschritte der Anfangsphase auch mit beiden erfolgt sein müssen. Allerdings nur bis zu dem Zeitpunkt, an dem das erste katalytisch wirkende Peptid entstand, das in der RNA gespeichert war und die weitere Entwicklung maßgeblich prägte (Start des Lebens).
Es sorgte dafür, dass durch die schnellere Bildung von Nachfolgemolekülen die Ressourcen stärker in dieser Gruppe (der heutigen D-Ribose und L-Aminosäuren) verbraucht wurden [18, 19]. Dass zu genau der gleichen Zeit ein derartiges Peptid sowohl in der D- als auch in der L-Version entstand, ist nahezu ausgeschlossen.
Eine Dreierbeziehung: Membran, Peptid und RNA - oder kurz, die MPR-Welt
Aus den bisherigen Überlegungen wird deutlich, dass es, wie oben beschrieben, keine isolierte RNA-, Peptid- oder Metabolismus zuerst- Welt gab. Vielmehr müssen drei Player gleichzeitig existiert haben, die eine chemische Evolution in gegenseitiger Abhängigkeit durchliefen: Die Membrane der Vesikel, durch die eine Trennung von hydrophoben und hydrophilen Aminosäuren möglich wurde, Peptide, die sich zu Beginn unspezifisch und danach informationsgesteuert bildeten und eine RNA, die aus einer geringstmöglichen Anzahl von Nukleotiden bestand. Abgekürzt wäre es die MPR-Welt.
Während in einigen anderen Modellen am Anfang eine Information für die Bildung von Enzymen gefordert wird, entstand nach dem hier vorgestellten Modell als erstes eine unspezifische Reihung von Codes, die mit einer Sequenz von Aminosäuren in Peptiden gekoppelt werden konnte. Erst, nachdem sich durch zufällige Kombinationen aus dem Pool der vielen unspezifischen Codes ein Proto-Enzym herauskristallisierte, das für die Weiterentwicklung der Zelle unterstützende Eigenschaften besaß, wurde die Information als solche in der RNA, die dadurch als Proto-mRNA etabliert wurde, festgelegt. Entsprechend wurde sie an die nachfolgenden Generationen durch Kopieren der Proto-mRNA weitergegeben. Erst durch die Bestätigung im Rahmen der Selektion bekam der in der RNA und später in der DNA gespeicherte Abschnitt seine Bedeutung.
Eine interessante Überlegung, die sich aus diesen Vorstellungen ergibt, ist die weitere Entwicklung der Enzymbildung. Durch sie wurde die Geschwindigkeit für die Entstehung der ersten Zellen vorgegeben.
Welche Situation ist denkbar? Es ist möglich, dass eine größere Anzahl von 12er Modulen zu einem längeren Proto-mRNA Strang verknüpft worden waren (Abb. 18-1.). Unter der Voraussetzung, dass spezifisch beladene Proto-tRNAs passende Plätze belegen und die transportierten Aminosäuren sich zu einer Kette verbinden konnten, war es nur eine Frage der Zeit, bis durch bestimmte Kombinationen katalytisch wirksame Peptide (Proto-Enzyme) entstanden.
Hierbei ist aber zu bedenken, dass immer nur Teile der Proto-mRNA belegt wurden, da noch nicht für alle Tripletts passende tRNAs und ihren Aminosäuren existierten und/oder nicht genügend Bausteine zur Verfügung standen. Ein Teil der heutigen kanonischen Aminosäuren traten erst später in der Entwicklung auf.
Man kann sich die Situation vorstellen wie ein Strang eines Keilriemens, dem etliche Zähne fehlen. Längere komplette Abschnitte sind durch Leerstellen begrenzt. Die längeren Abschnitte wären für die mRNA der Bereich, aus dem eine durchgehende Aminosäurekette gebildet werden konnte. Die begrenzenden Leerstellen hätten somit die Funktion der Start- und Stopp-Tripletts, die später für den Ableseprozess notwendig wurden.
Und jetzt wird es spannend: Wenn durch äußere Einflüsse oder durch inzwischen wirksame Enzyme eine neue Aminosäure mit zugehöriger tRNA (es gab genügend Auswahl, s.o.) im System erschien, konnten die passenden Tripletts der mRNA, die bisher frei geblieben waren, plötzlich belegt werden. Dies erfolgte nicht nur an einer Stelle, sondern über den gesamten Strang verteilt, immer dort, wo ein entsprechendes Codon vorlag. Im günstigsten Fall schloss sich so eine Lücke zwischen zwei Peptidkomplexen, die zusammen plötzlich eine höhere Funktionalität bekamen, als beide allein. Es bot die Chance, in kürzester Zeit neue große Peptide/Enzyme zu bilden, die wirksamere Katalysatoren darstellten, als die kleineren Ausgangspeptide für sich. Dies könnte in der Evolution jedes Mal einen großen Sprung nach vorn zur Folge gehabt haben. Ursache wäre im Vergleich zu einer Mutation, die nur punktuell an vielleicht einem Individuum wirkt, dass die Veränderung an vielen Stellen des Informationsspeichers greift, was gleichzeitig bei allen im Environment befindlichen RNAs möglich wurde.
Diese Vorstellung lässt sich gleich auch für die spätere Lebensentwicklung nutzen. Die nicht-kodierenden DNA-Abschnitte (sogenannte Junk-DNA) bieten viele Ansatzpunkte für die Etablierung neuer Enzyme. Sobald größere Umwälzungen im System Erde erfolgten, gab es schnelle Anpassungen der Lebewelt an die neuen Umweltbedingungen. Ein besonderer Einschnitt in der Erdgeschichte war zum Beispiel das Auftreten von Sauerstoff in der Atmosphäre. Durch Sauerstoff kann zum Beispiel die Nukleobase Guanin zu 8-Oxo-Guanin umgewandelt werden, das fehlerhaft mit Adenin statt Cytosin paaren kann. Es ist unklar, ob es diesen speziellen Effekt gab. Er soll nur zeigen, dass mit einem umweltbedingten Einfluss, der alle oder einen Großteil der Lebewesen betrifft, durch kleinste Änderungen in einem Bauteil der DNA, schnell deutliche Veränderungen in den Bauplänen der Enzyme u.a. entstehen können.

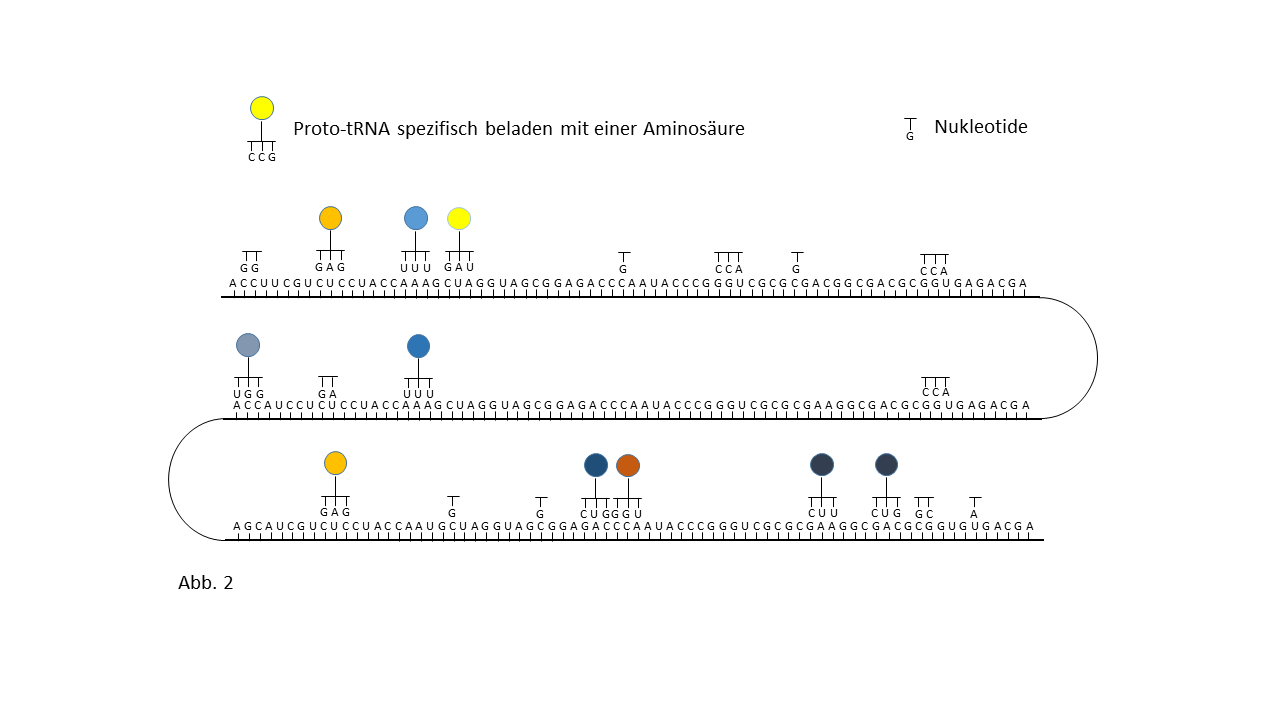
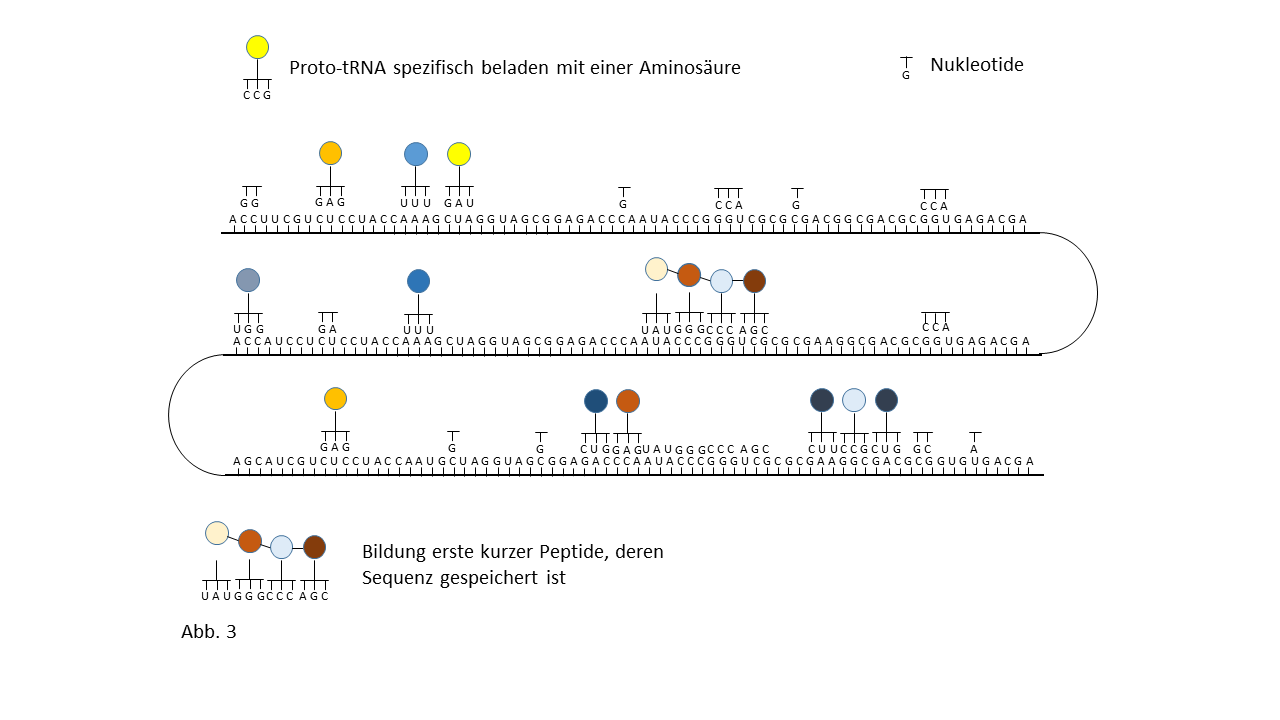
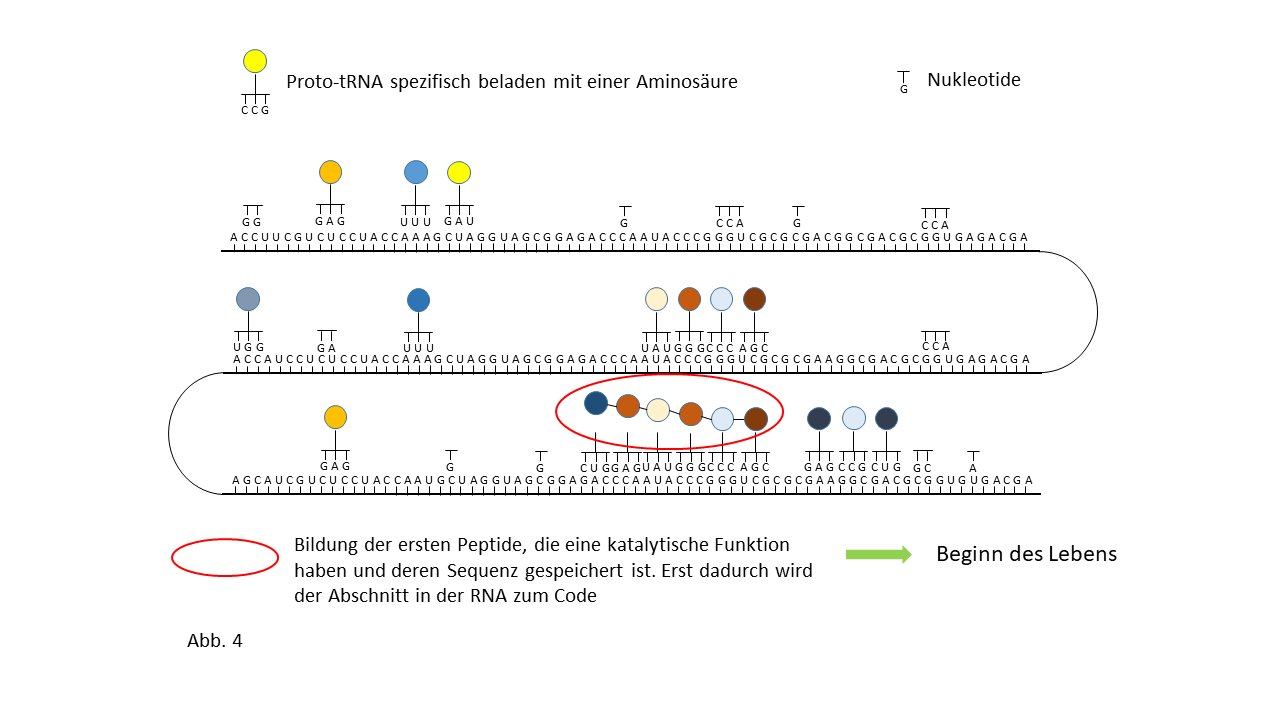
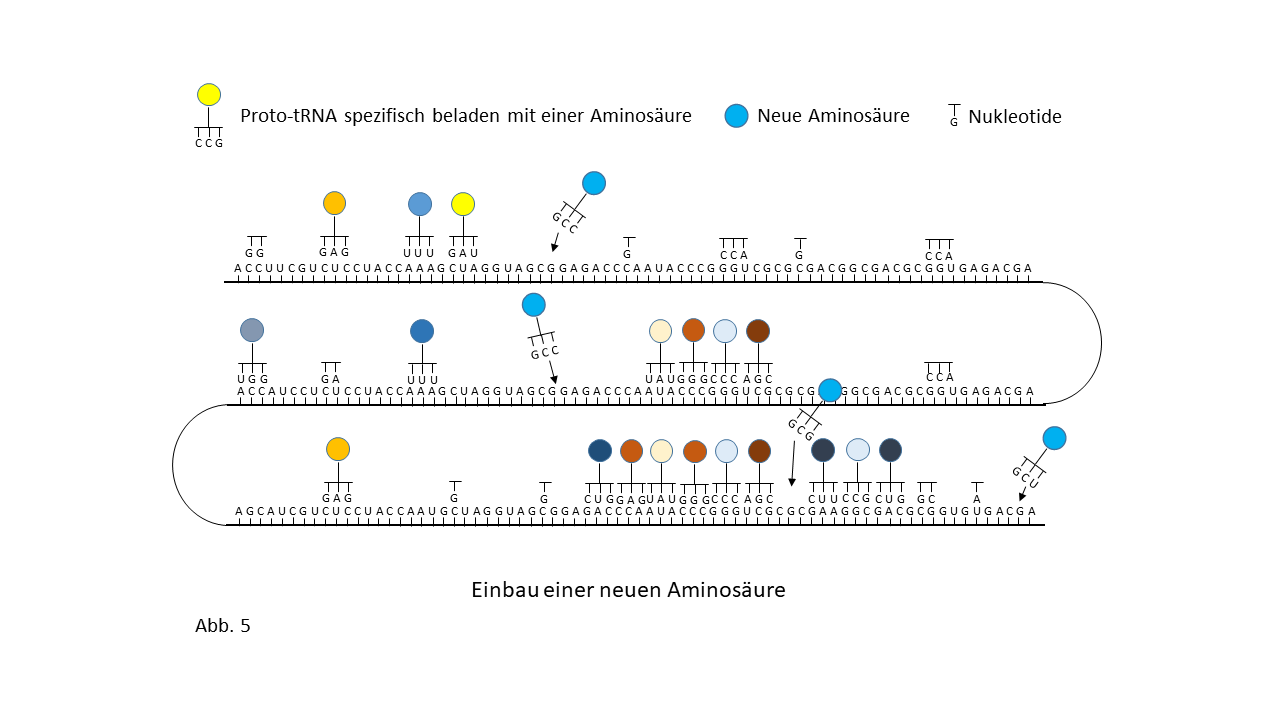
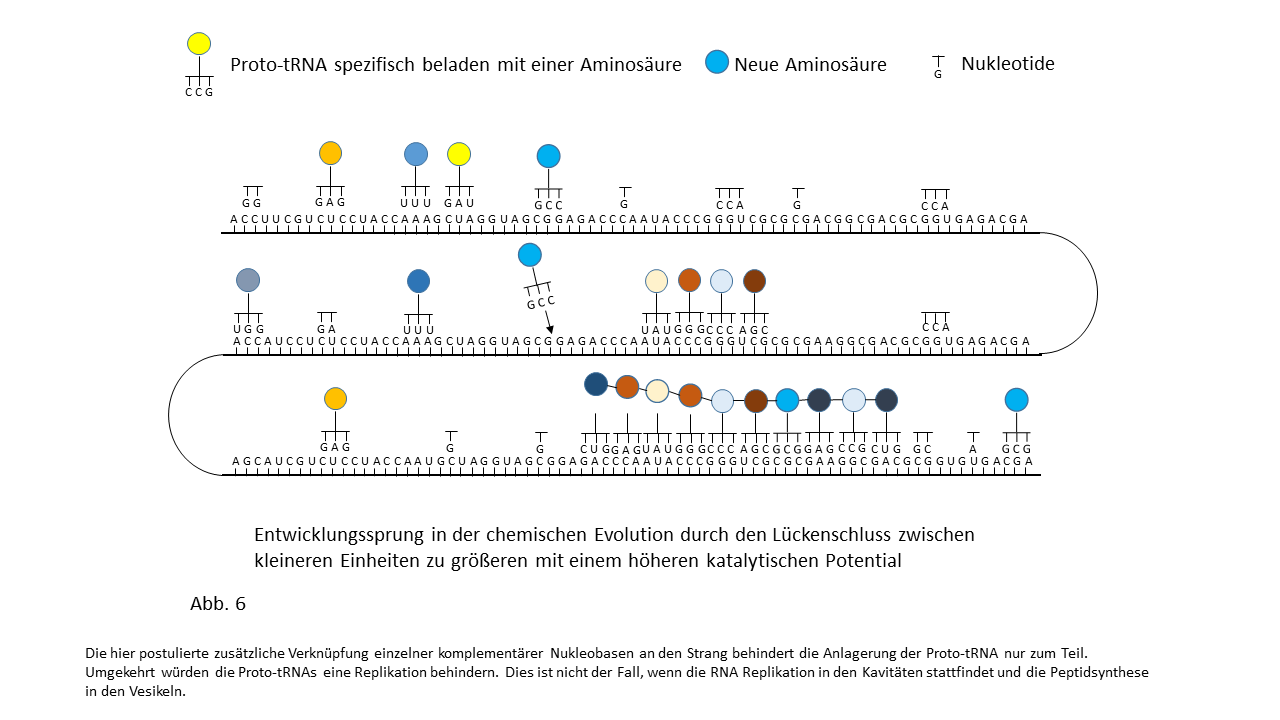
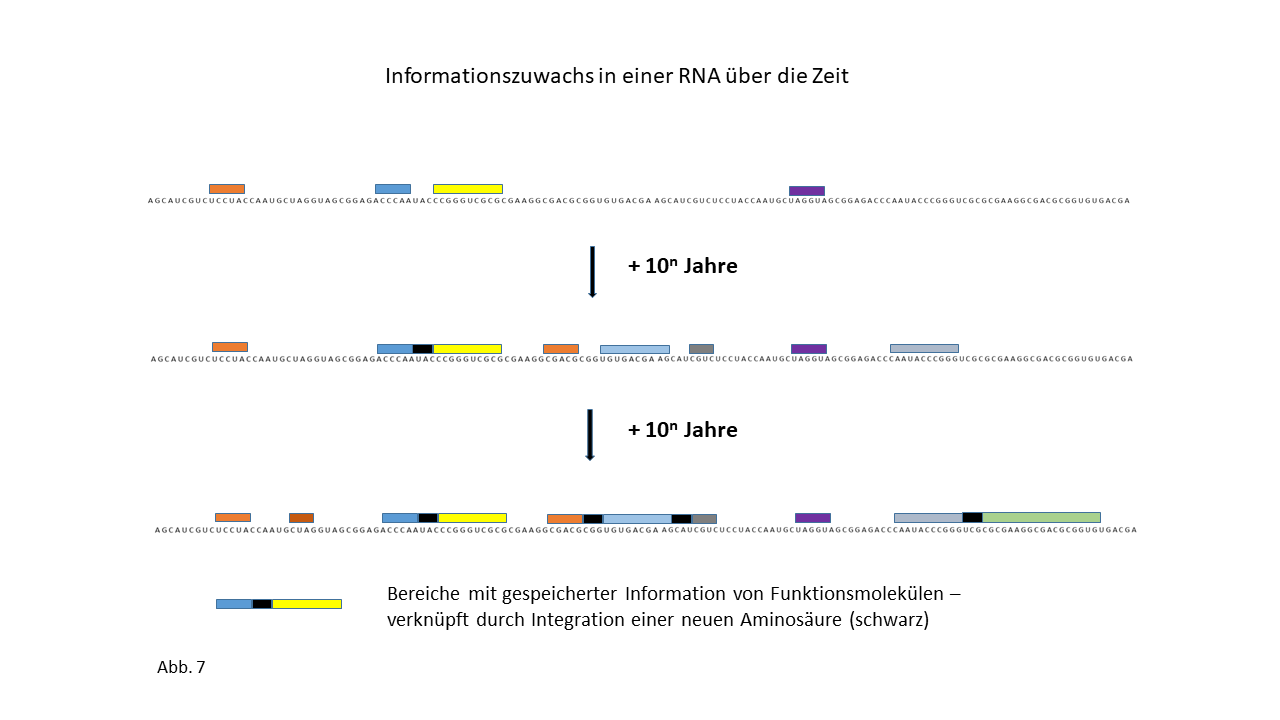
Abb. 18 1-7: Darstellung der Belegung eines längeren RNA-`Rohlings´ mit spezifisch beladenen Proto-tRNAs. Die Aminosäuren (farbige Punkte) sind über die Membranen der Vesikel mit den Proto-tRNAs verknüpft worden. (1) RNA-Rohling ohne Informationsinhalt und Funktion, der in den Kavitäten kopiert werden kann. (2) RNA-Rohling im Vesikel, wo eine statistische Belegung mit komplementären, spezifisch beladenen tRNAs erfolgt. (3) Bildung erster Peptide ohne Funktion. (4) Bildung erster Peptide, die im Verlauf der Evolution eine Funktion beinhalten und deren Sequenz gespeichert ist. Hiermit kann der Start des Lebens definiert werden. Die postulierte zusätzliche Verknüpfung einzelner komplementärer Nukleobasen an den Strang behindert die Anlagerung der Proto-tRNAs nur zum Teil. Umgekehrt würden die Proto-tRNAs eine Replikation behindern. Dies ist nicht der Fall, wenn die RNA Replikation in den Kavitäten und die Peptidsynthese in den Vesikeln stattfinden. (5) Integration einer neuen Aminosäure in das System. (6) Lückenschluss zwischen zwei Speicherabschnitten kleinerer Enzyme durch die neue Aminosäure zu einem neuen längeren mit effektiverer katalytischen Aktivität: Entwicklungssprung in der Evolution. (7) Zunahme der Funktionsabschnitte auf dem RNA Strang über die Zeit.
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Der Schlussstein
Es ist deutlich geworden, dass es ein Zusammenspiel von Vesikelbildungen und Prozessen in den Kavitäten der kontinentalen Bruchzonen gab. In den Kavitäten sammelten sich zuerst organische Moleküle, unter denen auch Bausteine für die Bildung der ersten Zellen waren. Mit der Vesikelbildung wurden diese Bausteine (Aminosäuren, Nukleobasen, Lipide u.a.) aufgenommen, wodurch sie im Inneren miteinander reagieren konnten [28].
Die Vesikel können als die Produktionsstätten der Enzyme angesehen werden, deren Information in den RNAs gespeichert wurde. Durch anschließenden Zerfall der Vesikel wurden die Moleküle in die Kavitäten freigegeben, sodass sie für den nächsten Zyklus wieder zur Verfügung standen. Es ist leicht einzusehen, dass die Lebensdauer der Vesikel von wenigen zehner Minuten nicht ausreichte, um komplexe Moleküle zu entwickeln. Auch die Entstehung uncodierter Peptide, die sich bei jedem Zyklus neu, mit einer zufälligen Sequenz bildeten und zu Membranporen zusammenschließen konnten, verlängerten die Lebensdauer der Vesikel nur unwesentlich.
Aber durch die Freisetzung der codierten Peptide mit dem Zerfall der Vesikel reicherten sich unterschiedlichste Enzyme über lange Zeiträume in den Kavitäten an. Auch wenn es Strömungen und Turbulenzen in den wassergefüllten Bruchzonen gab, waren genügend geschützte Bereiche vorhanden, in denen es zur Anreicherung kommen konnte. Mit der Zeit standen hierdurch immer mehr Funktionsmoleküle zur Verfügung, die mit jedem neuen Zyklus in die Vesikel aufgenommen werden konnten. Mit ihnen waren weitere, komplexe Reaktionen möglich. Aber am Ende des Zyklus wurde alles wieder in die Kavitäten überführt. Die Kavitäten entsprachen in diesem Zustand übergroßen Zellen (Giga-Zellen), die allerdings keine Membranumhüllung hatten.
Während die Speicherung der Information in den Vesikeln günstiger war, konnte die RNA eher in den Kavitäten kopiert werden (Abb. 19). Die hier vorliegende hohe Konzentration an Enzymen bei gleichzeitiger Existenz von RNA Strängen bot die ideale Voraussetzung für die Entwicklung komplexerer Molekül wie zum Beispiel das Ribosom oder die DNA. Die Lebensdauer des Environments war auf Millionen Jahre ausgelegt.
Das bedeutet, die Kavitäten waren gleichzeitig sowohl Reaktionsgefäße als auch Speicherorte für die Bausteine des Lebens. Mit jeder Vesikelbildung konnte aus dem Reservoir der Giga-Zelle geschöpft werden, sodass es nur eine Frage der Zeit war, bis eine günstige Mischung aus Funktionsmolekülen in ein oder mehrere Vesikel aufgenommen wurden, anhand derer der Start zur selbständigen Vermehrung vollzogen werden konnte (Abb. 20) [29].
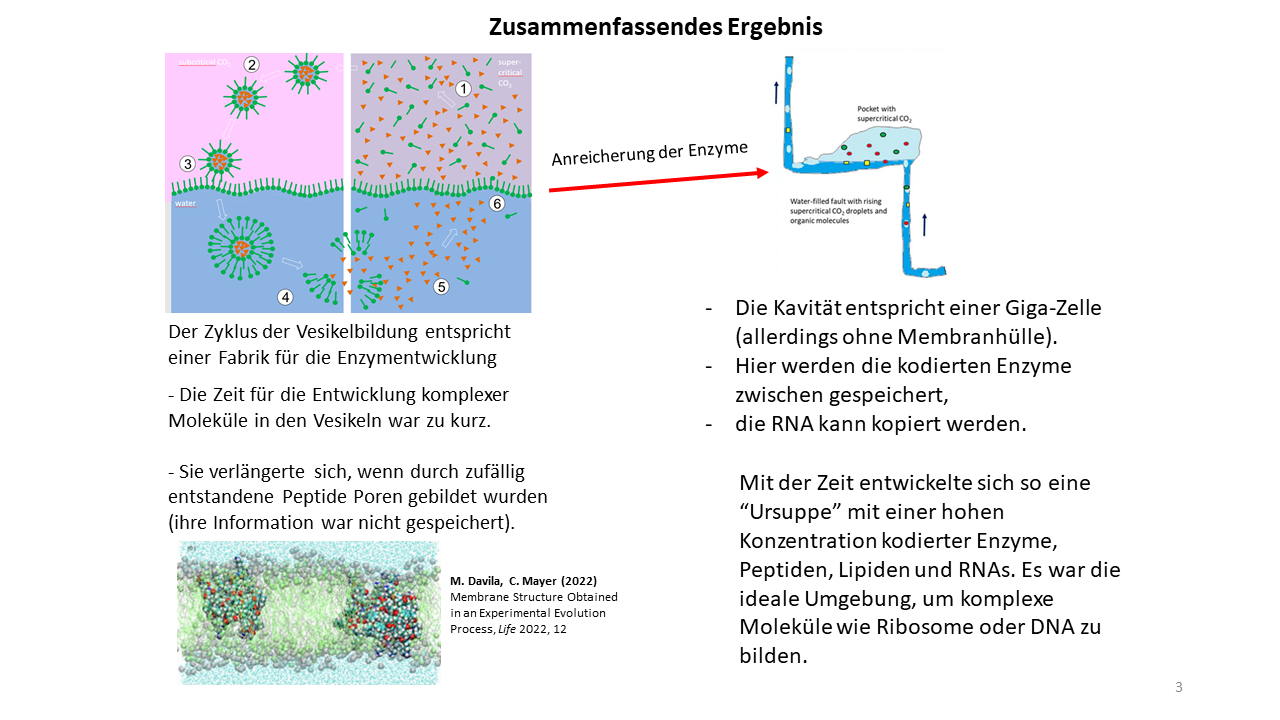
Abb. 19 Zusammenfassung der letzten Schritte auf dem Weg zur ersten Zelle
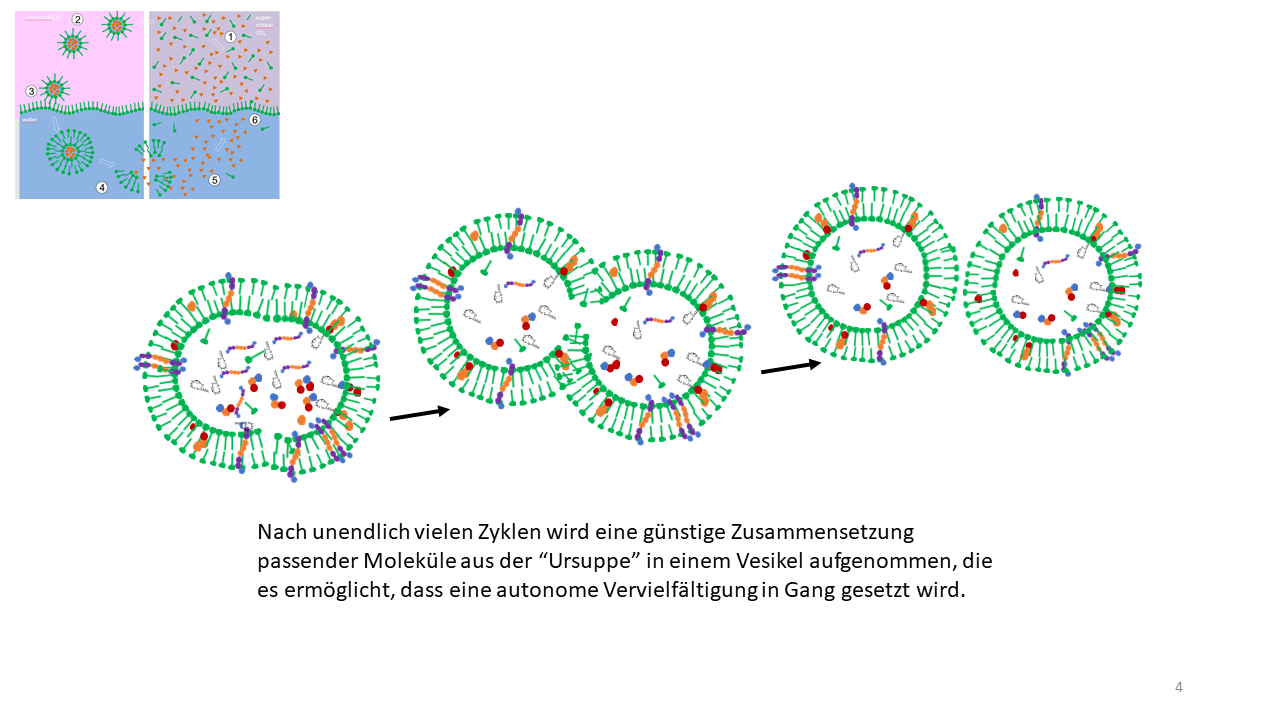
Abb. 20 Der entscheidende Zeitpunkt der optimalen Beladung mit Funktionsmolekülen eines Vesikels mit anschließender autonomen Vervielfältigung und physikalischen Teilung durch Scherung.
Mit dem Start der autonomen Vervielfältigung durch Stoffaufnahme von außen durch Poren und der katalytischen Steuerung der Molekülproduktion begann ein langer Zeitraum, in dem die internen Prozesse weiterentwickelt und optimiert wurden. Es dauerte vermutlich einige 100 Millionen Jahre, bis das System Zelle soweit entwickelt war, dass wir von LUCA Zellen sprechen können.
Und hier eine Alternative einer Proto-tRNA, die eine interessante mathematische Betrachtung möglich macht.
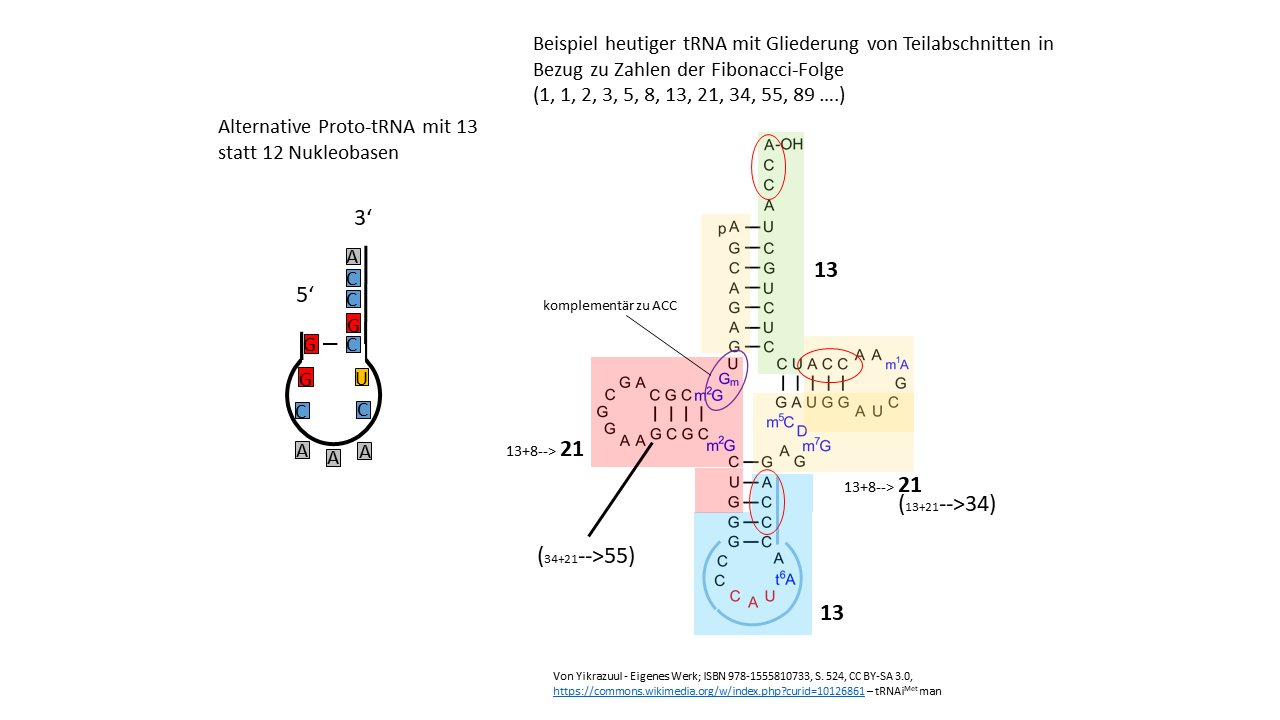
Wird die oben beschriebene Proto-tRNA um eine Nukleobase von 12 auf 13 verlängert, können die oben dargestellten Prozesse zur Informationsspeicherung, vergleichbar ablaufen. Es kann diskutiert werden, welche Proto-tRNA zu Beginn die wahrscheinlichste war, die 12er oder die 13er Version. 13 ist die siebte Zahl der Fibonacci-Folge, deren Zahlenspektrum häufig in der Natur zu finden ist.
Viele Pflanzenarten haben Blütenblätter, deren Anzahl einer Fibonacci-Zahl entspricht. Ebenso ist die Samenanordnung bei Sonnenblumen, Tannenzapfen oder Ananas in zwei Spiralrichtungen angelegt, deren Anzahl zwei aufeinanderfolgende Fibonacci-Zahlen sind. Die Schale des
Nautilus bildet eine Spirale, die durch das Aneinanderfügen von Quadraten mit den Seitenlängen der Fibonacci-Zahlen annähernd nachgebildet werden kann. Die Arme von Spiralgalaxien folgen oft einer logarithmischen Spirale, die sich aus der Fibonacci-Folge ableitet.
In der Abb. 20-1 ist links die alternative Proto-tRNA mit 13 Nukleobasen skizziert. Auf der rechten Seite steht die Kleeblattdarstellung für die tRNAMet des Menschen mit 74 Basen. Davon ausgehend, dass die Proto-tRNA mit 13 Nukleobasen die Grundlage für die Entwicklung der heutigen tRNA bildete, lässt sich der gesamte Strang in Einheiten untergliedern, deren Basenanzahl jeweils einer Zahl aus der Fibonacci-Folge entspricht.
Hiervon ausgehend kann überlegt werden, ob die nachfolgenden RNAs und anschließend die DNA aus Modulen aufgebaut wurden, die mit Zahlen aus der Fibonacci-Folge in Zusammenhang stehen. Jede vierte Zahl in der Fibonacci-Folge lässt sich durch 3 teilen. So zum Beispiel die 16. Zahl (987). Geteilt durch die 3 Basen des Codons sind das 329 Aminosäuren in einer Kette. Diese Zahl steht für gleich drei Enzyme/Proteine (rpoA, Humanes Cathepsin K (Präproprotein) und Protein 2C, ein hoch konserviertes und multifunktionales Protein bei Picornaviren, zu denen das Poliovirus gehört). Oder die 20. Zahl (6765) für 2255 Aminosäuren: Parainfluenza virus 5 (strain W3) (PIV5) (Simian virus 5).
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Abb. 20-1
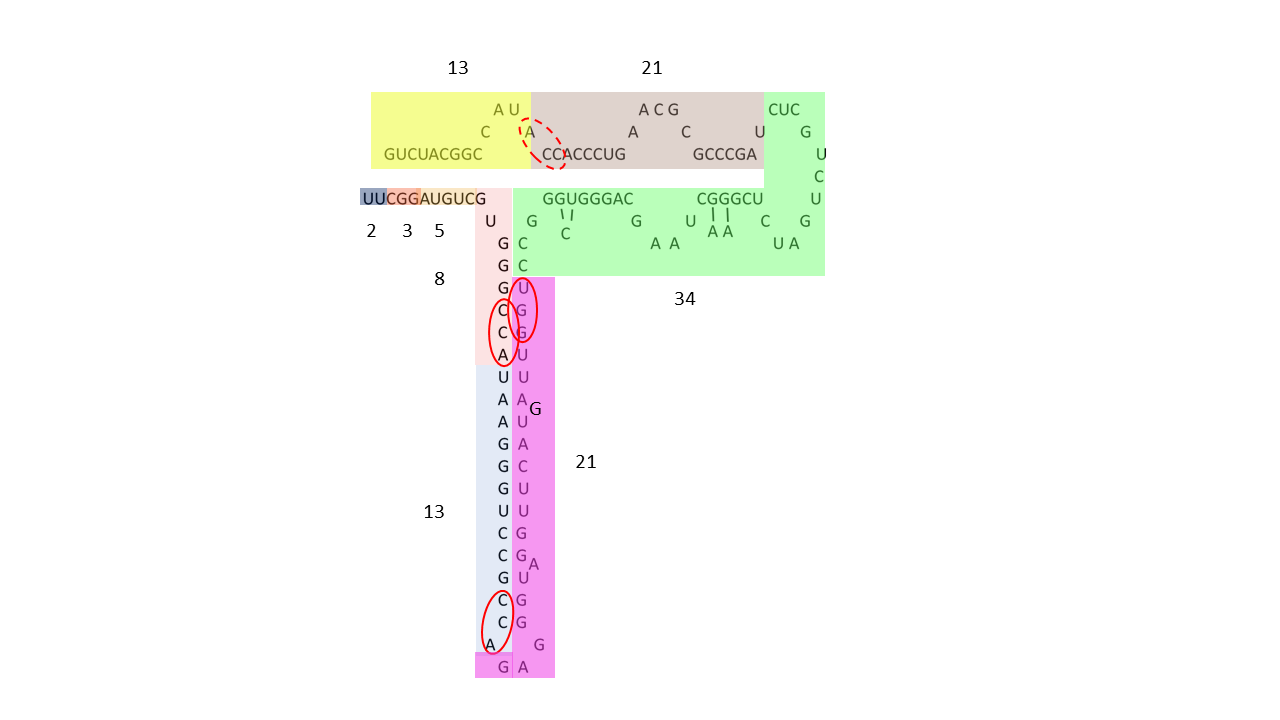
Die Einteilung der RNA nach Zahlen der Fibonacci-Folge ist bei völlig freiem Zugang hoch variabel und kann hinterfragt werden, da es eine Vielzahl von Kombinationsmöglichkeiten gibt. Wird aber vorausgesetzt, dass immer zwei beanchbarte Zahlen, wie z.B. 13 und 21 im Strang vorliegen müssen, sinkt die Anzahl der möglichen Kombinationen. Weiterhin kann aus dem oben dargestellten Proto-tRNA Modul mit 13 Nukleobasen häufig ACC oder das komplementäre Triplett UGG an den Anfang gestellt werden, wodurch die Richtung der Modulverknüpfung sichtbar wird. (Die gestrichelte rote Ellipse in Abb. 20-2 zeigt ein ACC Triplett, das möglicherweise am Anfang eines Moduls mit 21 Nukleobasen stand und durch späteren Umbau des Moleküls verschoben wurde.) Dies ist allerdings davon abhängig, ob die Sequenz hoch konserviert ist. Weiterhin ist durch die modulare Bauweise zu erwarten, dass mehrere Abschnitte, die bereits aus einer Kombiantion gebildet wurde, zu einer neuen, längeren Einheit verknüpft wurden. So wäre die heutige tRNA aus Abb. 20-1 aus zwei 13/21 Modulen zusammengesetzt.
Beispiel der 5S rRNA des Menschen.: 08.02.2026
Abb. 20-2: 5S rRNA aufgeteilt in Module aus Fibonacci-Zahlen. Ein Modul mit einer vollständigen Folge von 2 bis 34, kombiniert mit einem zweiten Modul mit 13/21; alternativ kann die 34 zu 13/21/34 zugeordnet werden. - rRNA Daten nach: Cheng, N., Mao, Y., Shi, Y., & Tao, S. (2012). Coevolution in RNA molecules driven by selective constraints: evidence from 5S rRNA.
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
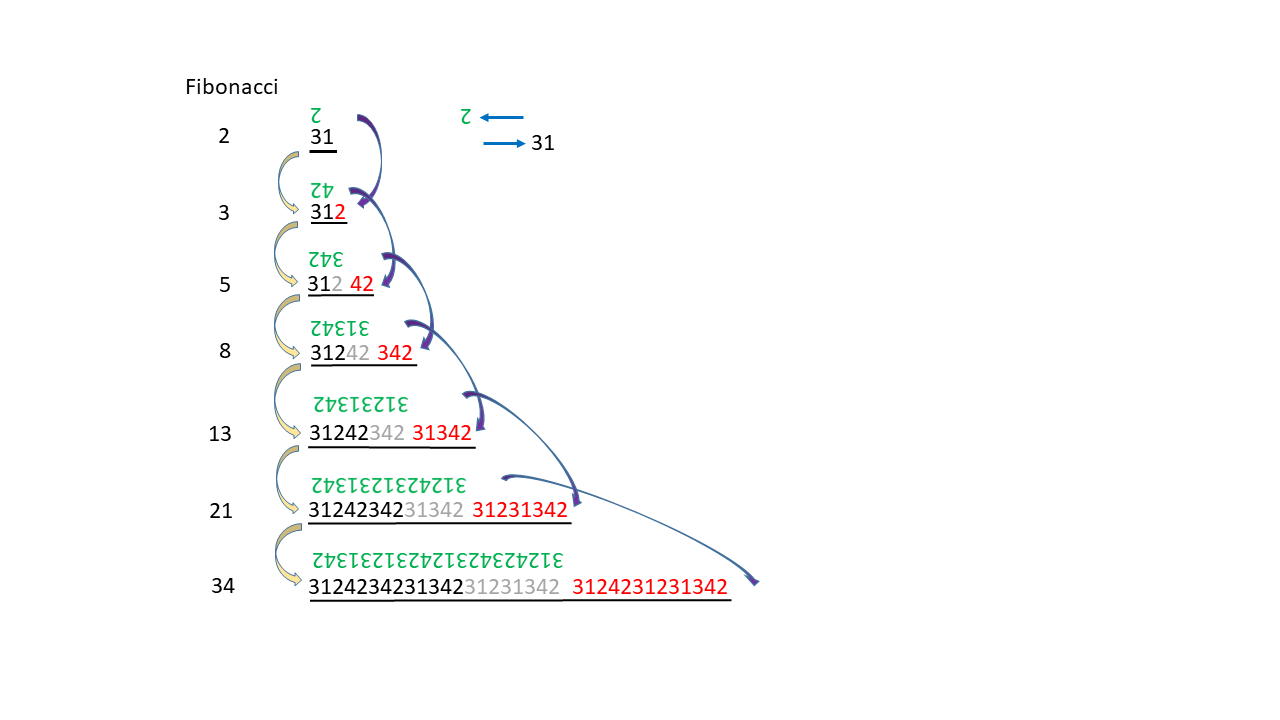
Einige unkonventionelle Gedanken (Spielereien) zu den Fibonacci Zahlen 04.04.2026
Wenn RNA-Strang Abschintte vielfach kopiert und die Kopien und Originale beliebig verknüpft werden, wiederholen sich in den länger werdenden Strängen immer die selben Abschnitte mit den selben Codes. Eine Zuordnung von Aminosäuren würde immer gleiche Sequenzen ergeben, wodurch die Variationsmöglichkeit der entstehenden Enzyme begrenzt ist. Eine hohe Variabilität in den Sequenzen ergibt sich dagegen, wenn die Verlängerung der RNA nicht immer in gleich langen Abschnitten erfolgt, sondern mit Zunahme von größer werdenden Teilstücken.
In der nachfogenden Abbildung ist ein Entwicklungsprozess skizziert, der die Verlängerung eines Zahlenstrangs (entstprechend einer RNA) zeigt, bei dem der jeweils nächste Schritt einer nächst höheren Fibonacci Zahl entspricht. Hierbei wird jeweils ein Teil des schwarzen Strangs kopiert (Kopie in grün). Die Zahlen von 1 bis 4 entsprechen den Nukleobasen A, G, U, und C. Die grüne Kopie ergänzt die jeweilige schwarze Zahl zur Summe 5. Die Ablese- bzw. Verknüpfungsrichtung ist mit den horizontalen Pfeilen angezeigt. Der Vorgang erfolgt folgendermaßen: Die Verlängerung des schwarzen Abschnitts (rote Zahlen) findet jeweils gleichzeitig mit der Kopie (grün) des bestehenden schwarzen Teils statt. Die rote Verlängerung stammt jeweils aus der grünen Kopie (dunkelblauer Pfeil beginnend bei den grünen auf dem Kopf stehende Zahlen). Die gelben Pfeile zeigen vom kleineren zum nächst größeren Strang. Die grauen Zahlen sind die vorherigen roten.
Vorteil könnte sein, dass längere Einzelstränge leicht einen Doppelstrang mit einem Loop in der Mitte bilden können.
Mirrkarioten
Der bisher aufgezeigte mögliche Weg zu den ersten Zellen trennt ab einem bestimmten Zeitpunkt die parallel verlaufende Entwicklung der Händigkeiten von L-Aminosäuren und D-Ribose der RNA der heutigen Welt von der alternativen Welt mit D-Aminosäuren und L-Ribose. Es stellt sich die Frage, ob die Entwicklung an allen der vielen möglichen Standorte gleich verlaufen ist, oder ob nach erfolgreicher Ausbreitung der ersten Zellen von einem Ursprungsort aus, die lokalen Verhältnisse an anderen Stellen soweit überprägt wurden, dass sie fortan auch in die gleiche Richtung liefen. Der letzte Fall erscheint am wahrscheinlichsten.
Es gibt aber vermutlich eine gewisse Chance, dass an einem Standort, vielleicht auf einem anderen Kleinkontinent, eine ähnliche Entwicklung zur ersten Zelle ablief, wie beschrieben, die aber zum entgegengesetzten, spiegelbildlichen System führte. Dies wäre neben der uns bislang bekannten Lebensform LF1 eine zweite Form, die LF2. Während die LF1 in die Domänen der Bakterien und Archaeen (beides Prokarioten) sowie Eukarioten unterteilt werden, könnte die LF2 aus nur einer Domäne bestanden haben. Für sie wäre die Bezeichnung Mirrkarioten passend. Bei diesen Überlegungen taucht sofort die Frage auf, ob es diese „Mirrkarioten“ heute noch geben könnte, in unzugänglichen Bereichen wie der tiefen Biosphäre, vielleicht in den ältesten Kontinentkernen.
Und daran schließt sich gleich die wichtigste Frage an. Würden wir sie überhaupt mit unseren analytischen Methoden entdecken und von den anderen Einzellern unterscheiden können? Vielleicht leben sie auch schon immer unter uns, ohne dass wir von ihnen Notiz nehmen konnten.
Wie ging es weiter? - Die nächsten Schritte
Die Geysir-Eruptionen beförderten kontinuierlich organische Moleküle und schließlich die ersten selbstvermehrenden Zellen an die Erdoberfläche. Im Umfeld der Austrittsstellen müssen sich folglich „Bio-Filme“ aus organischen Materialien gebildet haben, in die auch die ersten Zellen eingebettet wurden. Sie hatten allerdings unter den neuen Umweltbedingungen kaum Überlebenschancen. UV-Strahlung, Sonnenwind, niedrigere Temperaturen und höhere pH-Werte erforderten eine Anpassung, die einen längeren Zeitraum benötigte. Durch rücklaufendes Wasser in die Störungszonen gelangten die angepassten Moleküle und ersten Zellen wieder in den Tiefenprozess der Kruste.
Darüber hinaus war die Energie- und Stoffzufuhr schwierig. Der Biofilm im Umfeld des Geysirs bot aber vermutlich die Voraussetzung, dass die Zellen Stoffe von außen aufnehmen konnten. Früher oder später können klimatische oder regionale Bedingungen dafür gesorgt haben, dass ein Überleben der Zellen auf der Erdoberfläche möglich wurde, sodass sie über Fließgewässer in die Ozeane gelangen konnten. Eine alternative Situation ist für küstennahe Standorte denkbar, deren offene Störungssysteme direkt in den marinen Raum überleiteten. Der Eintritt in den Ozean war der entscheidende Schritt, der den Siegeszug des Systems Leben in Bewegung setzte. Im Ozean trafen die Zellen auf die Gebiete, die ihnen von den physikochemischen Verhältnissen vertraut waren. Es waren die Black Smoker und andere hydrothermale Quellen, die ihnen unbegrenzte, nutzbare Energieressourcen bereitstellten. Gleichzeitig bot dieses Umfeld erneut Schutz vor zerstörerischen Einflüssen, die an der Erdoberfläche vorhanden waren.
Die Inbesitznahme der submarinen hydrothermalen Quellen durch die ersten prokariotischen Zellen führte, zeitlich komprimiert betrachtet, vermutlich zu einer explosionsartigen Ausbreitung der ersten Lebensform. Der offene Ozean bot mit Meeresströmungen und ausreichend vielen heißen Quellen hierfür ideale Bedingungen. Große Distanzen und lokale Besonderheiten konnten zu eigenständigen Anpassungen in der jungen Familie der Zellen führen, die somit schnell diversifizierten. In dieser Phase lag das Ausklingen der heftigen Meteoriteneinschläge, die überwiegend die Ozeane betrafen. Mit jedem größeren Einschlag wurden weite Bereiche der jungen Kontinente überflutet. Dies war der Weg, auf dem sich die Prokaryoten über alle großen und kleinen Krustenfragmente ausbreiten konnten. Überall dort, wo Kontakt zu hydrothermalen Quellen an Land bestand, konnten sie Fuß fassen und sich weiterentwickeln. In diesem Zusammenhang ist eine weitere Entwicklung auf der jungen Erde von Bedeutung. Schon früh setzte die Bewegung von Platten ein, die letztlich zur Verlagerung der Kontinente und zu nachfolgenden Kollisionen führte. Welchen Einfluss hatte dieser Prozess auf die weitere Entwicklung der Prokarionten?
Inzwischen gilt es als gesichert, dass Organellen wie die Mitochondrien und Chloroplasten der Eukaryoten von Bakterien abstammen. Es besteht die Vorstellung, dass Bakterien von Archaeen aufgenommen wurden, aber nicht verdaut werden konnten. Im Gegenteil, die aufgenommenen Bakterien lebten in der Zellhülle des Wirtes weiter und nutzten deren Stoffwechselprodukte für die eigene Versorgung. Es entwickelte sich eine Endosymbiose, bei der der eine Partner in dem Körper des anderen dauerhaft lebt, wie die Bakterien im Darm. Die evolutive Anpassung führte dazu, dass die „unverdauten“ Bakterien nach und nach zu nützlichen Bausteinen reduziert wurden, die heute für die Energieversorgung der Zelle einen entscheidenden Beitrag leisten.
Die Eukarioten traten vor 1,5 Milliarden Jahren fast spontan auf der Bühne des Lebens auf. Vorfahren, die als Übergangsstadien angesehen werden könnten, sind bisher nicht eindeutig identifiziert. 500 Millionen Jahre vorher begannen fast alle bestehenden Kleinkontinente aufeinander zuzuwandern, bis sich 300 Millionen Jahre später ein Superkontinent gebildet hatte. Er wird von den Geologen als Columbia bezeichnet. Sein Bestehen reichte über weitere 200 – 300 Millionen Jahre, wobei die letzten 100 Millionen Jahre bereits vom Zerfall und plattentektonisch bedingten Gebirgsbildungsprozessen geprägt waren. In der Endphase von Columbia traten schließlich die Eukarioten auf. Bereits eine Milliarde Jahre vorher begann die Sauerstoffkonzentration in der Atmosphäre langsam zuzunehmen, auf den die Lebewelt reagieren musste.
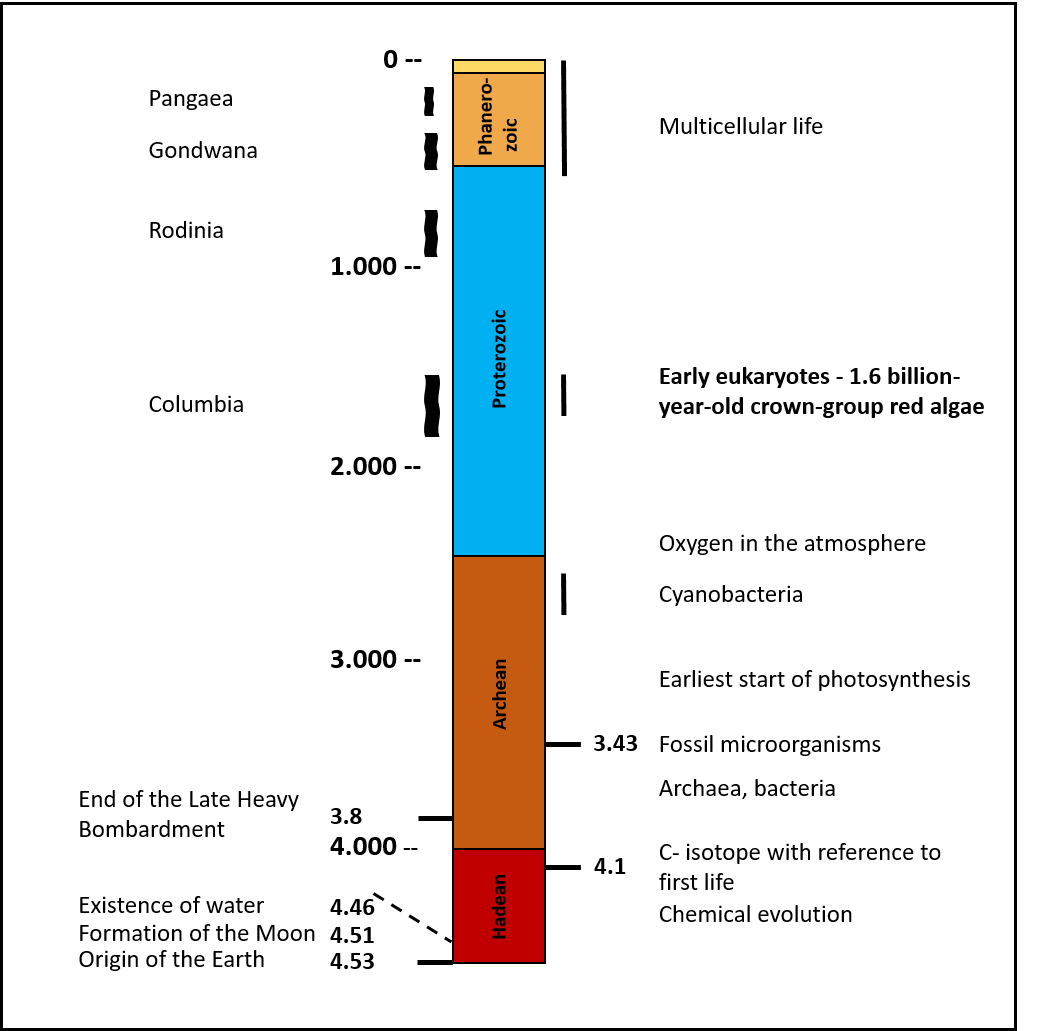
Abb. 21 Zeitstrahl der Erdgeschichte - Alter der Eukaryoten nach Bengtson et al. 2017 [21]
Angenommen auf allen weit auseinanderliegenden Kleinkontinenten hatten sich über mehr als eine Milliarden Jahre endemische Bakterienstämme entwickelt, die ab einem bestimmten Zeitpunkt innerhalb von Millionen Jahren nach und nach in einem Großkontinent zusammengeführt wurden. Jedes Mal, wenn ein neuer Kleinkontinent an dem großen Komplex anlandete, standen sich „plötzlich“ unterschiedlich stark spezialisierte Archaeen und Bakterien gegenüber. Die Anzahl der Individuen und der Zeitraum waren groß genug, dass alle Variationen des Zusammenlebens ausprobiert werden konnten. Es ist denkbar, dass dieser Prozess zur Entwicklung der Endosymbionten führte, vielleicht durch nachfolgend auftreffende Kleinkontinente sogar mehrfach, wie es bei einigen Organellen der Zellen (Plastiden der Braun-, Gold- und Kieselalgen) heute beobachtet werden kann. Wenn hierzu parallel die Anpassung an die größer werdenden Sauerstoffkonzentrationen in der Atmosphäre und dem Ozeanwasser verlief, könnte alles zusammen genommen am Ende von Columbia den
Start der Eukarioten erklären.
Aus diesem hypothetischen Scenario lassen sich gleich einige Fragen anschließen. Gibt es heute in der
DNA der Bakterien und Archaeen noch Relikte, die auf die Zeit der endemischen Entwicklung hinweisen? Lassen sich vielleicht Provinzen auf den Kontinenten finden, die alten Kontinentkernen entsprechen und wo derartige Bakterien und Archaeen vorkommen, vielleicht in der
tiefen Biosphäre?
Reflexionen
Die oben vorgestellte Theorie zur Entstehung des Lebens verleitet dazu, einmal über den Tellerrand zu schauen, um mögliche weiterreichende Zusammenhänge zu erkennen. Es führt zwangsläufig zu einer mehr philosophischen Betrachtungsweise, die die bisherigen Erkenntnisse aus einer anderen Perspektive in Beziehung setzen.
Das, was als Kern des Modells heraussticht, ist das Bild einer größeren Reaktionskammer, in der Informationen in Form von codierten Molekülen kontinuierlich gespeichert werden. Innerhalb dieses Raumes entstehen bei wiederkehrenden Abläufen mit der Bildung von Vesikeln wesentlich kleinere Einheiten, die die Informationen (Moleküle) aufnehmen, verändern und nach Zerfall der Kompartimente wieder in die große Kammer abgeben. Dies geschieht in unendlich vielen Vorgängen, bis sich einige der Vesikel aufgrund ausreichend aufgenommener Information selbständig weiterentwickeln können.
Diese Bild erinnert an die getrennt geschlechtliche Vermehrung, bei der, z.B. beim Menschen, aus einer großen Anzahl von Informationsträgern ein Träger selektiert wird, der die Information in einem kleinen Kompartiment, dem Ei, nach vielen Versuchen zu einer vermehrungsfähigen Einheit vervollständigt.
Ein Schritt dazwischen vollzog sich mit dem Übergang in der Zellentwicklung von den Prokarioten zu den Eukarioten, der sich ca. eine Milliarde Jahre nach dem Auftreten des ersten Lebens vollzog. Hierbei entstand aus Zellen der Archaeen Linie durch Aufnahme von Bausteinen/Informationen aus einem anderen Einzellertyp nach unendlich vielen Vorgängen eine neue Lebensform. Die Bausteine stammten von Bakterien, die sich separat in verschiedenen großen Environments entwickelt hatten. Mit den Eukarioten begann eine neue Lebensgemeinschaft, die komplexer war, als das Leben vorher.
Wenn wir in die heutige Entwicklung blicken, sehen wir die Sammlung von Information aus einem großen vom Menschen geschaffenen Wissenspool. Sie wird durch Verknüpfung und Verdichtung zu einer neuen, komplexen Einheit entwickelt, an deren Ende eine KI, ein Programm existiert, das in der Lage sein wird, sich materiefrei im Universum zu verbreiten.
Ist diese Betrachtung auf die Zeit des uns bekannten Lebens begrenzt?
Wir haben mit der Bildung der Planeten und des Sonnensystems eine Konzentration von Materie an einem spezifischen Ort. Die Materie enthält Informationen, die bei der Kondensation aus einer viel größeren Einheit, dem kosmischem Nebel und Galaxien, zusammengetragen wurde. Hierbei gab es Phasen unterschiedlicher Einträge, die zu der heutigen Zusammensetzung der Erde führten. Über die Entwicklungszeit des Weltalls haben in unzähligen Regionen ähnliche Bedingungen geherrscht, wie sie zur Bildung der Erde geführt haben. Aus der Kenntnis der Gesetzmäßigkeiten des irdischen Lebens lässt sich folgern, dass eine Vielzahl von vergleichbaren Entwicklungen im Universum gestartet wurden, von denen wahrscheinlich ist, dass eine gewisse Anzahl ebenfalls (zumindest einfaches) Leben hervorbrachten.
Und an dieser Stelle sei noch ein Blick auf das Gesamtsystem erlaubt.
Unser Universum schein nach Ansicht der meisten Physiker ein besonderer Glücksfall zu sein, bei dem die fundamentalen Nuturkonstanten genau die Größen haben, bei denen sich Materie, die physikalischen Prozesse und Leben entwickeln konnten. Schon eine geringe Abweichung bei einigen Konstanten würde ein Universum hervorbringen, in dem komplexe Strukturen und somit auch das Leben nicht entstehen könnten (Stärke der Gravitationskraft, der elektromagnetischen Kraft, der starken Kernkraft, der schwachen Kernkraft; die Größe der kosmologischen Konstante, die Masse von Elektronen und Quarks).
Mit dem Modell der Entstehung des Lebens vor Augen lässt sich überlegen, ob es nicht genau so viele Versuche wie bei der Vesikelbildung gegeben hat, um aus einer gigantisch großen Einheit kosmische Blasen zu erzeugen, die sich jedes Mal mit einem Urknall zu einem Universum entwickelten. Wie bei den Vesikel könnte es hierbei überwiegend Fehlversuche gegeben haben, weil die Naturkonstanten nicht die optimalen Werte besaßen. In wenigen Fällen, oder nur in einem war es entsprechend erfolgreich. Dies würde dem von Physikern diskutierten Modell des Multiversums entsprechen. Das Weltall driftet auseinander, und wie bei den Vesikeln, wird es zerfallen und vielleicht wieder in der gigantisch großen Ausgangseinheit aufgenommen.
Fundraising
zur Unterstützung meiner Forschung zur Entstehung des Lebens und angrenzenden Wissenschaftsgebieten
Die Forschung zur Entstehung des Lebens gehört zu einem extrem interdisziplinären Bereich, der bisher nicht von dritter Seite gefördert wurde. Es sind keine ausgetretenen Pfade, die beschritten werden konnten, es geht um etwas völlig Neues, etwas das neue Einsichten über unsere Welt erkennen lässt. Es liefert letztendlich die Antworten, die wir suchen, um verstehen zu können, wo wir herkommen, wie es weitergehen kann und ob wir vielleicht nicht allein im Universum sind. Darüber hinaus wird sich mit dem Wissen um die Anfänge die medizinische Forschung gezielter weiterentwickeln, soviel ist sicher.
Meine bisherige Forschung beruht bis heute auf viel Eigeninitiative und dem Einsatz privater Mittel. Um sie fortzusetzen (ich bin bereits im Ruhestand) ist eine finanzielle Unterstützung sehr hilfreich. Die Gelder sollen für internationale/nationale Tagungen, Projektunterstützungen, Fahrtkosten ins Gelände, Veröffentlichungskosten und andere Kosten verwendet werden. Hierzu gehört unter anderem die Weiterführung des Youtube Kanals `Exciting Geology´, über den Forschungsergebnisse der breiten Öffentlichkeit präsentiert werden.
Auf Wunsch werden die Namen der Spender aufgeführt. Eine Kommunikation über die Verwendung der Spenden ist selbstverständlich.
Kontoinhaber: Prof. Dr. Ulrich Schreiber
Das Spendenkonto (ING Bank): IBAN DE39 5001 0517 5822 6603 00 BIC: INGDDEFFXXX
Stichwort: Forschungsspende
Literatur
[1] Schreiber, U., Locker-Grütjen, O., & Mayer, C. (2012). Hypothesis: Origin of life in deep-reaching tectonic faults. Origins of Life and Evolution of Biospheres, 42(1), 47-54.
[2] Mayer, C., Schreiber, U., & Dávila, M. J. (2015). Periodic vesicle formation in tectonic fault zones—an ideal scenario for molecular evolution. Origins of Life and Evolution of Biospheres, 45(1), 139-148.
[3] Dhuime, B., Hawkesworth, C. J., Cawood, P. A., & Storey, C. D. (2012). A change in the geodynamics of continental growth 3 billion years ago. Science, 335(6074), 1334-1336.
[4] Mayer, C., Schreiber, U., Dávila, M. J., Schmitz, O. J., Bronja, A., Meyer, M., Klein, J. & Meckelmann, S. W. (2018). Molecular evolution in a peptide-vesicle system. Life, 8(2), 16.
[5] Schreiber, U., Mayer, C., Schmitz, O. J., Rosendahl, P., Bronja, A., Greule, M., Keppler, F., Mulder, I., Sattler, T. & Schöler, H. F. (2017). Organic compounds in fluid inclusions of Archean quartz—Analogues of prebiotic chemistry on early Earth. PLoS One, 12(6), e0177570.
[6] Großmann, Y., Schreiber, U., Mayer, C., & Schmitz, O. J. (2022). Aliphatic Aldehydes in the Earth’s Crust—Remains of Prebiotic Chemistry?. Life, 12(7), 925.
[7] Benner, S. A., Kim, H. J., & Carrigan, M. A. (2012). Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Accounts of chemical research, 45(12), 2025-2034.
[8] Pedreira-Segade, U., Feuillie, C., Pelletier, M., Michot, L. J., & Daniel, I. (2016). Adsorption of nucleotides onto ferromagnesian phyllosilicates: Significance for the origin of life. Geochimica et Cosmochimica Acta, 176, 81-95.
[9] Cleaves II, H. J., Scott, A. M., Hill, F. C., Leszczynski, J., Sahai, N., & Hazen, R. (2012). Mineral–organic interfacial processes: potential roles in the origins of life. Chemical Society Reviews, 41(16), 5502-5525.
[10] Shih, P., Pedersen, L. G., Gibbs, P. R., & Wolfenden, R. (1998). Hydrophobicities of the nucleic acid bases: distribution coefficients from water to cyclohexane. Journal of molecular biology, 280(3), 421-430.
[11] Jarvinen, P., Oivanen, M., & Lonnberg, H. (1991). Interconversion and phosphoester hydrolysis of 2', 5'-and 3', 5'-dinucleoside monophosphates: kinetics and mechanisms. The Journal of Organic Chemistry, 56(18), 5396-5401.
[12] Mayer, C., Schreiber, U., & Dávila, M. J. (2017). Selection of prebiotic molecules in amphiphilic environments. Life, 7(1), 3.
[13] Eriani, G., Delarue, M., Poch, O., Gangloff, J., & Moras, D. (1990). Partition of tRNA synthetases into two classes based on mutually exclusive sets of sequence motifs. Nature, 347(6289), 203-206.
[14] Baillet, J., Desvergnes, V., Hamoud, A., Latxague, L., & Barthélémy, P. (2018). Lipid and nucleic acid chemistries: combining the best of both worlds to construct advanced biomaterials. Advanced Materials, 30(11), 1705078.
[15] Marshall, W. L. (1994). Hydrothermal synthesis of amino acids. Geochimica et Cosmochimica Acta, 58(9), 2099-2106.
[16] Hennet, R. C., Holm, N. G., & Engel, M. H. (1992). Abiotic synthesis of amino acids under hydrothermal conditions and the origin of life: a perpetual phenomenon?. Naturwissenschaften, 79(8), 361-365.
[17] Weber, A. L., & Lacey, J. C. (1978). Genetic code correlations: amino acids and their anticodon nucleotides. Journal of Molecular Evolution, 11(3), 199-210.
[18] Schreiber, U. C. (2019). Das Geheimnis um die erste Zelle: dem Ursprung des Lebens auf der Spur. Springer-Verlag.
[19] Schreiber, U. C., & Mayer, C. (2020). The first cell. Springer International Publishing.
[20] Kyte, J., & Doolittle, R. F. (1982). A simple method for displaying the hydropathic character of a protein. J Mol Biol, 157(1):105-132.
[21] Bengtson, S., Sallstedt, T., Belivanova, V., & Whitehouse, M. (2017). Three-dimensional preservation of cellular and subcellular structures suggests 1.6 billion-year-old crown-group red algae. PLoS Biology, 15(3), e2000735.
[22] Jungck, J. R. (1978). The genetic code as a periodic table. Journal of Molecular Evolution, 11, 211-224.
[23] Tamura, K., Schimmel, P. (2004). Chiral-selective aminoacylation of an RNA minihelix. Science, 305, 1253.
[24] Suga, K., Tomita, H., Tanaka, S., & Umakoshi, H. (2012). Hydrophobic properties of tRNA with varied conformations evaluated by an aqueous two-phase system. International Journal of Biological Sciences, 8(8), 1188.
[25] Carter, C. W. (2017). Coding of Class I and II aminoacyl-tRNA synthetases. Protein Reviews: Volume 18, 103-148.
[26] Czerniak T, Saenz JP (2025) The secret life of RNA and lipids. RNA Biology, VOL. 22, NO. 1, 1–28.
https://doi.org/10.1080/15476286.2025.2526903)
[27] Michanek A, Yanez M, Wacklin H, Hughes A, Nylander T, Sparr E (2012) RNA and DNA association to zwitterionic and charged monolayers at the air–liquid interface. Langmuir, 28(25), 9621-9633.
[28] Dávila, M. J., & Mayer, C. (2023). Structural phenomena in a vesicle membrane obtained through an evolution experiment: A study based on MD simulations. Life, 13(8), 1735.
[29] Schreiber, U. (2025). The Origin of Life in the Early Continental Crust: A Comprehensive Model. Life, 15(3), 433.